Advances in Animal and Veterinary Sciences
Short Communication
Advances in Animal and Veterinary Sciences. 1 (1S): 14 – 17Special issue–1 (Veterinarians approaches for safeguarding animal health and production)
Pre–Partum Serum Cytokine Levels as a Potential tool for the Prediction of Retention of Fetal Membranes in Cross–Bred Cows
Jones Boby, Harendra Kumar*, Ajevar Ganesan, Sanjay Kumar Singh, Krishnaswamy Narayanan
-
Animal Reproduction Division, Indian Veterinary Research Institute, Izatnagar–243122 India
*Corresponding author:hkumar1960@gmail.com
ARTICLE CITATION:
Jones Boby J, Kumar H, Ganesan A, Singh SK and Narayanan K (2013). Pre–partum serum cytokine levels as a potential tool for the prediction of retention of fetal membranes in cross–bred cows. Adv. Anim. Vet. Sci. 1 (1S): 14 – 17.
Received: 2013–08–20, Revised: 2013–08–30, Accepted: 2013–08–30
The electronic version of this article is the complete one and can be found online at
(
http://nexusacademicpublishers.com/table_contents_detail/4/98/html
)
which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited
ABSTRACT
India, by virtue of its huge cattle population ranks second in world cow milk production, however the per–capita availability of milk is less owing to the lower production potential of cows. This could be attributed to the harsh tropical climate and high incidence of reproductive disorders. Among the various postpartum reproductive disorders, the incidence of retention of fetal membranes (RFM) is highest. Identifying the RFM cows ante–partum would help in the better management and minimize its adverse consequences on the post–partum fertility. During the peri–partum period, a cytokine mediated immune mechanism operates at foeto–maternal interface for the successful placental separation and any failure or delay in this process precipitates RFM. Hence, we hypothesized that the estimation of serum level of cytokines– IL–10, TNF–α and IL–6 would, in part, reveal the immune changes taking place during advanced pregnancy and aid in predicting the RFM. Accordingly, the levels of IL–10, TNF–α and IL–6 were estimated in pre–partum serum samples of 81 cows by bovine specific Enzyme linked immunoabsorbant assay (ELISA). Out of 81 cows studied, 8 developed RFM and the rest calved (n=73) normally. IL–10 and TNF–α concentration was found to be significantly higher in RFM cows than their normal counterparts (P<0.01), whereas the IL–6 concentration was higher (P<0.01) in normal cows than the RFM group. This study for the first time demonstrates that these cytokines are associated with the development of RFM and, therefore, their significant level of detection could serve as a predictive tool. However, studies with large number of RFM cows are needed to ascertain the utility of these cytokines as markers.
India is home for 15% of world cattle population and ranks second in the world’s cow milk production (FAOSTAT 2010). Despite the overall higher milk production, the productivity is lower owing to the harsh tropical climate and higher incidence of reproductive disorders. Retention of fetal membranes (RFM has been found to be the single largest postpartum complication in the bovine species and is defined as the failure to expel placenta even after 12–24 hrs of calving (Leblanc, 2008; Mohamed and Amer, 2009); whereas, in majority of normal cows (66%), placental expulsion occurs in within 6 hrs of calving. The incidence of RFM ranged from 4.0 to16.1% in different herds in various countries (Sheldon 2008; Mohamed and Amer 2009); whereas in India, it ranged from 4.5 to 28.2% (Dongre et al., 2011). RFM has a multifactorial etiology (Hanafi et al., 2011) and it adversely affects the fertility by delaying the uterine involution and increasing the time to first service, services per conception and days open as the condition causes metritis and endometritis (McDougall et al., 2001). RFM cows are also at high risk of suffering from ketosis and mastitis (Beagley et al., 2010). Therefore, any tool in predicting the occurrence of RFM may help in initiating prophylactic measures, thus preventing and alleviating its negative consequences on fertility.
During pregnancy, the maternal immune response towards the fetus bearing paternal allo–antigens are modulated rather than suppressed for a successful pregnancy outcome. There is partial down regulation of the expression of MHC antigens on placenta (Low et al., 1990); production of immuno–suppressant molecules like Interferon– τ (Thatcher et al., 1986; Spencer et al., 2007) and also a shift from Th1 to Th2 type immune responses (McCracken et al., 2004). Th2 cells promote antibody responses which are more conducive for pregnancy and produce anti–inflammatory cytokines–IL–10, IL–4 and IL–5 inhibiting the induction of cytotoxic Th1 response. In the second and third trimesters of healthy pregnancy, Th2 cells predominate in the peripheral blood (Saito et al., 1999). Reversal of Th2 toTh1 shift during the peripartum period is essential to cope up with the imminent microbial contamination of the postpartum uterus (McCracken et al., 2004; Sheldon, 2009). Any delay in this shift usually predisposes the cow for postpartum reproductive disorders.
Cytokines exert antiluteolytic, antiviral, antimicrobial and immunomodulatory effects during pregnancy, and also in the regulation of embryo growth and differentiation (Schafer–Somi 2003). The MHC class I expression on fetal trophoblast cells are unregulated during third trimester of pregnancy and initiates an inflammatory reaction by way of cytokine synthesis, eventually resulting in placental expulsion (Davies et al., 2000, 2004). Altogether, the increased pro–inflammatory cytokine levels have a significant role in placental separation, increasing the collagenase activity that culminates in the dehiscence of foetal cotyledon from the maternal caruncle (Beagley et al., 2010).
TNF–α and IL–6 are the pro–inflammatory cytokines with authoritative role in wide range of inflammatory, infectious, autoimmune and malignant conditions; produced in response to infection primarily by monocytes/macrophages and T and B lymphocytes (Apostolaki et al., 2010; Scheller et al., 2011). Sustained levels of increased TNF–α in the serum could reflect persistent uterine bacterial infection. IL–6 operates by two signaling pathways out of which classic signaling mediates regenerative and anti–inflammatory activities whereas trans–signaling brings about pro–inflammatory responses (Scheller et al., 2011). IL–10 is a general suppressive cytokine which inhibits pro–inflammatory responses from innate and adaptive immunity and has a key role in the resolution of inflammation (Filippi et al., 2008). This study was aimed to predict RFM cows by determining the prepartum serum levels of pro and anti–inflammatoty cytokines during peri–partum period.
A total of 81 healthy advanced pregnant cows between 2nd and 4th parity which had no history of reproductive abnormalities during the previous gestation were selected, randomly at 240 days of pregnancy, from a herd of cross bred cows maintained at an organized dairy farm, IVRI, Izatnagar. The animals were maintained under uniform feeding and managemental conditions. The pregnancy diagnosis was performed routinely in the above mentioned farm at 50–60 days post insemination. The expected date of calving was fixed at 280 days post insemination based on artificial insemination data supported with record of pregnancy diagnosis. Five ml of blood was collected from the jugular vein at 15 days before the expected date of calving using a vacutainer and the serum was separated.. The serum was aliqouted and stored at –20oC till analysis for assay of cytokines– IL–10, TNF–α and IL–6.
The cows were closely observed on the day of calving for the occurrence of any complications and the time of expulsion of placenta was recorded. The cows which expelled placenta within 24 hrs of calving were categorized as Non–RFM (n=73) and those which retained placenta for more than 24 hrs were grouped as RFM (n=8).The serum concentration of IL–10, TNF–α and IL–6 was estimated using commercially available bovine specific ELISA kits (Blue Gene, India). The ELISA kit works on the principle of competitive enzyme immunoassay technique, making use of a monoclonal anti–cytokine antibody and a cytokine–HRP conjugate.
All the kit components and samples were brought to room temperature (20–25oC ) before start of assay and performed as per manufacturer’s instructions. The optical density was measured immediately at 450 nm using a microplate reader. A standard curve was obtained by plotting the concentration of the standards (0, 50, 100, 250, 500 and 1000 pg/ml) against their optical densities. The value of unknown samples was interpolated from the standard curve and was presented in pg/ml.
Statistical analysis was performed using SPSS software version 16. Independent sample t– test was used to compare the values between cows with and without RFM.
The IL–10 concentration was found to be significantly (P<0.001) higher in RFM cows than those without RFM (1798.5 ± 143.44 Vs 944.28 ± 129.25 pg/ml). Similarly, the cows with RFM had a significantly higher (P<0.01) levels of TNF–α (911.35±64.69 pg/ml) as compared to those which did not experience RFM (675.54±38.33 pg/ml). However, a lower serum level of IL–6 was observed in RFM (1139.4 ± 95.90 pg/ml) than Non–RFM cows (1657.2 ± 77.977 pg/ml) (Table1).
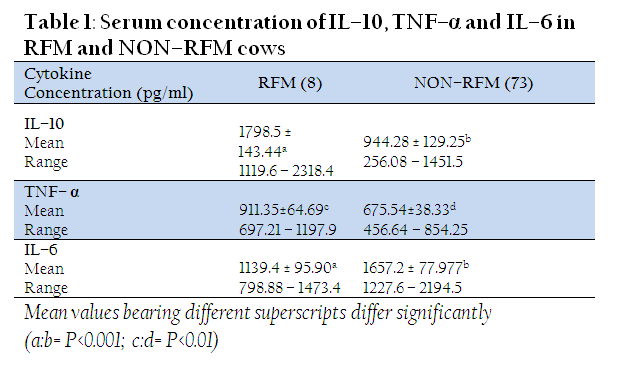
The higher IL–10 levels in RFM cows observed in the present study are well supported by Islam et al. (2013) who recently reported a significantly higher IL–10 level in pre–partum serum samples (15 days before expected calving) of crossbred cows which subsequently developed RFM and clinical metritis (CM). They also reported a higher serum IL–10 level on the day of calving in cows with RFM and CM as compared to normally calved cows. IL–10 being a potent anti–inflammatory cytokine is capable of suppressing pro–inflammatory cytokine production and hinders class II MHC and CD14 expression, thereby inhibiting T cell activation by macrophages (Ouyang et al., 2011). The main biological activity of IL–10 is inhibiting the production of monocyte/macrophage derived TNF–α, IL–6, IL–1, IL–8, IL–12, Granulocyte colony stimulating factor (GCSF) and also cytokine production by neutrophils and natural killer cells (Opal and Depalo, 2000). It is known that anti–inflammatory cytokines are produced in concurrence with pro–inflammatory cytokines and their levels indicate the severity of the preceding infection (Couper et al., 2008).
The exact cause responsible for this pre–partum rise in IL–10 levels in cows which subsequently developed RFM is still not clear. The higher IL–10 levels might have suppressed the T cell activation and macrophage activity, thus inhibiting the mechanisms responsible for placental separation. The placental separation, an immune mechanism by itself, might have failed to occur in presence of a strong anti–inflammatory environment created by IL–10.
Further, the significantly higher levels of TNF–α in RFM cows are also in agreement with Islam (2012) who observed a similar pattern of TNF–α in cows with RFM and CM at approximately 15 days before the due calving. Experiments in murine models have detected a rise in the plasma levels of pro–inflammatory cytokines viz., TNF–α, IL–6 and IL–1β even in normal pregnancy during late gestation and assumed it as a normal regulatory or immune mechanism taking place during the periparturient period (Orsi et al., 2006). A higher concentration of TNF–α was observed after lipopolysaccharide (LPS) injection in ewes (Dow et al., 2010), LPS induced abortion in cows (Foley et al., 1993) and also in LPS stimulation of whole blood from cows (Rontved et al., 2005).
TNF–α is a potent pleiotropic pro–inflammatory cytokine with definitive role in inflammatory, infectious, autoimmune and some malignant conditions (Apostolaki et al., 2010). It is the end product of NFkB signaling pathway which is initiated by binding of pathogen associated molecular patterns (PAMP) on microbial organisms to their respective pattern recognition receptors on host cells (Fitzgerald et al., 2000; Bryant et al., 2010). How this pathway gets activated during the periparturient period, especially in the RFM cows, is yet to be determined. Bacterial contamination of the immune suppressed uterus or whether any genetic predisposition is behind it needs to be ascertained. The placental retention occurring even in the high TNF–α milieu suggest that this cytokine may only be a contributory factor in placental separation rather than having a direct effect.
The significantly higher serum levels of IL–6 in normally– calved cows than those of RFM cows are also in the concurrence with the findings of Ishikawa et al. (2004) who reported lower prepartum serum IL–6 concentrations in cows which subsequently developed RFM. They also reported a higher prepartum IL–6 levels compared to post–partum. The low level of IL–6 in RFM cows would have compromised the reaction to the allograft (placenta) and eventually resulted in RFM (Ishikawa et al., 2004). Further, IL–6 levels in cervico–vaginal fluid of women and in dried blood spot specimens from preterm neonates were found to be increased during spontaneous preterm birth, when compared to full–term infants (Wei et al., 2010). Serum IL–6 level monitoring proved useful for detecting the acute rejection of hepatic allografts in studies conducted with monkeys and higher peaks was observed in recipients who rejected grafts (Ohzato et al. 1993). All these studies suggest the important role of IL–6 in maintenance of pregnancy and the initiation of labour.
Thus it can be opined that higher serum levels of anti–inflammatory cytokine IL–10 in RFM cows would have contributed to the decreased levels of IL–6 in the present study. IL–6 being a pro–inflammatory cytokine is an essential component of immune mechanisms and a fall in its level might have derailed the normal immune mediated separation taking place at the foeto–maternal interface and precipitated RFM.
A significantly higher pre–partum serum concentration of IL–10 and TNF–α was observed in cows which developed RFM, whereas IL–6 concentration was lower in RFM cows than those of normally calved ones. This suggested that an immune mechanism involving cytokines is of importance in placental separation. The present study demonstrated that these cytokines are associated with the development of RFM and, therefore, could serve as a predictive tool. However, further studies with large number screening of RFM cows is needed to ascertain their utility as markers.
ACKNOWLEDGEMENT
This study is part of the MVSc research work conducted by the first author. The authors thank ICAR and Director IVRI for the financial assistance.
CONFLICT OF INTEREST
The authors have no conflict of interest to declare.
REFERENCES
Apostolaki M, Armaka M, Victoratos P and Kollias G (2010). Cellular mechanisms of TNF function in models of inflammation and autoimmunity. Curr. Dir. Autoimmun. 11: 1 – 26.
http://dx.doi.org/10.1159/000289195
PMid:20173385
Beagley JC, Whitman KJ, Baptiste KE and Scherzer J (2010). Physiology and treatment of retained fetal membranes in cattle. J. Vet. Int. Med. 24: 261 – 268.
http://dx.doi.org/10.1111/j.1939-1676.2010.0473.x
PMid:20136715
Bryant D, Becker L and Richardson J (2010). Cardiac failure in transgenic mice with myocardial expression of tumor necrosis factor–α. Circulation 97: 1375 – 1381.
http://dx.doi.org/10.1161/01.CIR.97.14.1375
Couper KN, Blount DG and Riley EM (2008). IL–10: The master regulator of immunity to infection. J. Immunol. 180: 5771 – 5777.
http://dx.doi.org/10.4049/jimmunol.180.9.5771
PMid:18424693
Davies CJ, Fisher PJ and Schlafer DH (2000). Temporal and regional regulation of major histocompatibility complex class I expression at the bovine uterine/placental interface. Placenta 21: 194 – 202.
http://dx.doi.org/10.1053/plac.1999.0475
PMid:10736242
Davies CJ, Hill JR, Edwards JL, Fisher PJ and Schlafer DH (2004). Major histocompatibility antigen expression on the bovine placenta: Its relationship to abnormal pregnancies and retained placenta. Anim. Reprod. Sci. 82–83: 267 – 280.
http://dx.doi.org/10.1016/j.anireprosci.2004.05.016
PMid:15271459
Dongre VB, Balasundaram B and Gupta AK (2011). Incidence of post parturient utero–vaginal complications in dairy cattle: a review. Wayamba J. Anim. Sci. 15: Number 1295082777.
Dow TL, Rogers–Nieman G, Holaskova I, Elsasser TH and Dailey RA (2010). Tumor necrosis factor– and acute–phase proteins in early pregnant ewes after challenge with peptidoglycan–polysaccharide. Domest. Anim. Endocrinol. 39: 147 – 154.
http://dx.doi.org/10.1016/j.domaniend.2010.04.001
PMid:20603027
Filippi CM and Von Herrath MG (2008). IL–10 and the resolution of infections. J. Pathol. 214: 224 – 230.
http://dx.doi.org/10.1002/path.2272
PMid:18161757
Fitzgerald KA, O’Neill LAJ and Gearing AJH (2001). The Cytokine Facts Book, First edition, Academic Press, San Diego.
Foley N, Lambert C, Mc Nicol M, Johnson N and Rook GAW (1990). An inhibitor of the toxicity of tumor necrosis factor in the serum of patients with sarcoidosis, tuberculosis and Crohn's disease. Clin. Exp. Immunol. 80: 395 – 399.
http://dx.doi.org/10.1111/j.1365-2249.1990.tb03299.x
PMid:1695561 PMCid:PMC1535197
Hanafi EM, Ahmed WM, El–Khadrawy HH and Zabaal MM (2011). An overview on placental retention in farm Animals. Middle–East J. Sci. Res. 7: 643 – 651.
Ishikawa Y, Nakada K, Hagiwara K, Kirisawa R, Iwai H, Moriyoshi M and Sawamukai, Y (2004). Changes in interleukin–6 concentration in peripheral blood of pre– and post–partum dairy cattle and its relationship to postpartum reproductive diseases. J. Vet. Med. Sci. 66: 1403 – 1408.
http://dx.doi.org/10.1292/jvms.66.1403
PMid:15585955
Islam R (2012) Studies on immune–endocrine profile of peripartum cows in relation to postpartum reproductive health. Thesis submitted to IVRI, Izatnagar, India
Islam R, Kumar H, Nandi S and Rai RB (2013). Determination of anti–inflammatory cytokine in periparturient cows for prediction of postpartum reproductive diseases. Theriogenology 79: 974 – 979.
http://dx.doi.org/10.1016/j.theriogenology.2013.01.018
PMid:23434356
LeBlanc S (2008). Postpartum uterine disease and dairy herd reproductive performance – A review. Vet. J. 176: 102 – 114.
http://dx.doi.org/10.1016/j.tvjl.2007.12.019
PMid:18328749
Low BG, Hansen PJ and Gogolin–Ewens KJ (1990). Expression of major histocompatibility complex antigens on the bovine placenta. J. Reprod. Fertil. 90: 235 – 243.
http://dx.doi.org/10.1530/jrf.0.0900235
PMid:2231545
McCracken SA, Gallery E and Morris JM (2004). Pregnancy–specific down regulation of NF–kappa B expression in T cells in humans is essential for the maintenance of the cytokine profile required for pregnancy success. J. Immunol. 172: 4583 – 4591.
http://dx.doi.org/10.4049/jimmunol.172.7.4583
PMid:15034076
McDougall S (2001). Effects of periparturient diseases and conditions on the reproductive performance of New Zealand dairy cows. NZ Vet. J. 49: 60 – 68.
http://dx.doi.org/10.1080/00480169.2001.36204
http://dx.doi.org/10.1080/00480169.2001.36223
Mohamed A and Amer A (2009). Hormonal and biochemical anomalies in dairy cows affected by retained fetal membranes. http://priory.com/vet/cow_fetal_membrane.htm.
Ohzato H, Monden M, Yoshizaki K, Gotoh M, Kanai T, Umeshita K, Tono T, Nishimoto N, Kishimoto T and Mori T (1993). Serum interleukin–6 levels as an indicator of acute rejection after liver transplantation in cynomologous monkeys. Surgery Today 23: 521 – 527.
http://dx.doi.org/10.1007/BF00730628
PMid:7689373
Opal SM and Depalo VA (2000). Anti–inflammatory cytokines. Chest 117: 1162 – 1172.
http://dx.doi.org/10.1378/chest.117.4.1162
PMid:10767254
Orsi NM, Gopichandran N, Ekbote UV and Walke JJ (2006). Murine serum cytokines throughout the estrous cycle, pregnancy and postpartum period. Anim. Reprod. Sci. 96: 54 – 65.
http://dx.doi.org/10.1016/j.anireprosci.2005.11.010
PMid:16413151
Ouyang W, Rutz S, Crellin NK, Valdez A and Hymowitz SG (2011). Regulation and functions of the IL–10 family of cytokines in inflammation and disease. Ann. Rev. Immunol. 29: 71 – 109.
http://dx.doi.org/10.1146/annurev-immunol-031210-101312
PMid:21166540
Rontved CM, Anderson JB, Dernfalk J and Ingvartsen KL (2005). Effects of diet energy density and milking frequency in early lactation on tumour necrosis factor–alpha responsiveness in dairy cows, Vet. Immunol. Immunopathol. 104: 171 – 181.
http://dx.doi.org/10.1016/j.vetimm.2004.11.001
PMid:15734538
Saito S, Sakai M, Sasaki Y, Tanebe K, Tsuda H and Michimata T (1999). Quantitative analysis of peripheral blood Th0, Th1, Th2 and the Th1: Th2 cell ratio during normal human pregnancy and preeclampsia. Clin. Exp.Immunol. 117: 50 – 555.
http://dx.doi.org/10.1046/j.1365-2249.1999.00997.x
PMCid:PMC1905376
Schafer–Somi S (2003). Cytokines during early pregnancy of mammals: a review. Anim. Reprod. Sci. 75; 73 – 94.
http://dx.doi.org/10.1016/S0378-4320(02)00222-1
Scheller J, Chalaris A, Schmidt–Arras D and Rose–John, S (2011). The pro– and anti–inflammatory properties of the cytokine interleukin–6. Biochimica et Biophysica Acta 1813: 878 – 888.
http://dx.doi.org/10.1016/j.bbamcr.2011.01.034
PMid:21296109
Sheldon IM, Williams EJ, Miller AN, Nash DM and Herath S (2008). Uterine diseases in cattle after parturition. Vet. J. 17: 115 – 121.
http://dx.doi.org/10.1016/j.tvjl.2007.12.031
PMid:18329302 PMCid:PMC2706386
Sheldon IM, Cronin J, Goetze L, Donofrio G and Schuberth HJ (2009). Defining postpartum uterine disease and the mechanisms of infection and immunity in the female reproductive tract in cattle. Biol. Reprod. 81: 1025 – 1032.
http://dx.doi.org/10.1095/biolreprod.109.077370
PMid:19439727 PMCid:PMC2784443
Spencer TH, Ott TL and Bazer FW (1996). τ–Interferon: pregnancy recognition signal in ruminants. Proc. Soc. Exp. Biol. Med. 213: 215 – 229.
http://dx.doi.org/10.3181/00379727-213-44053
PMid:8985305
Thatcher WW, Bazer FW, Sharp DC and Roberts RM (1986). Interrelationships between uterus and conceptus to maintain corpus luteum function in early pregnancy: sheep, cattle, pigs and horses. J. Anim. Sci. 62: 25 – 46.
PMid:3533874
Wei SQ, Fraser W and Luo ZC (2010). Inflammatory cytokines and spontaneous preterm birth in asymptomatic women: A systematic review. Obstet. Gynec. 116: 393 – 401
http://dx.doi.org/10.1097/AOG.0b013e3181e6dbc0
PMid:20664401



