Advances in Animal and Veterinary Sciences
Research Article
Advances in Animal and Veterinary Sciences 1 (4): 111 – 115Epidemiology and Genetic Diversity of Rotavirus Strains Associated with Acute Gastroenteritis in Bovine, Porcine, Poultry and Human Population of Madhya Pradesh, Central India, 2004–2008
Yashpal Singh Malik1*, Naveen Kumar1, Kuldeep Sharma1, Rajeev Sharma2, Harsh Bardhan Kumar2, Kusumakar Anupamlal2, Savita Kumari3, Sanjay Shukla4, KM Chandrashekar2
- Indian Veterinary Research Institute, Izatnagar 243122, Uttar Pradesh, India
- JN Krishi Vishwa Vidyalaya, Jabalpur 482001, Madhya Pradesh, India
- Department of Microbiology, Bihar Veterinary College, Patna, India
- College of Veterinary Science and Animal Husbandry, Madhya Pradesh Pashu Chikitsha Vigyan Viswavidyalaya, Rewa, M.P., India
*Corresponding author:malikyps@ivri.res.in
ARTICLE CITATION:
Malik YS, Kumar N, Sharma K, Sharma R, Kumar HB, Anupamlal K, Kumari S, Shukla S and Chandrahekar KM (2013). Epidemiology and genetic diversity of rotavirus strains associated with acute gastroenteritis in bovine, porcine, poultry and human population of Madhya Pradesh, Central India, 2004–2008. Adv. Anim. Vet. Sci. 1 (4): 111 – 115.
Received: 2013–08–17, Revised: 2013–09–05, Accepted: 2013–09–05
The electronic version of this article is the complete one and can be found online at
(
http://www.nexusacademicpublishers.com/table_contents_detail/4/80/html
)
which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited
ABSTRACT
Virus induced enteritis is one of the grave problems accounting for maximum deaths in neonatal animals and human throughout the world. In developing countries like India, the socio–economic conditions favors the occurrence of higher disease frequency of gastro–enteric infections, majorly dominated by rotavirus (RV). The dearth of appropriate surveillance programs and laboratory facilities have resulted in scarcity of accessible data on RV associated enteric disease affliction and epidemiological evidence in the country. We describe here the epidemiological and genotypic distribution of RV in various animal species and human during 2004 to 2008 from Madhya Pradesh, the central region of India. The overall prevalence rate of RV was 17.19% (126/733), maximum in humans (30.23%), followed by buffaloes (22.01%), cattle (13.33%), swine (13.04%) and poultry (6.47%). The nested–polymerase chain reaction (PCR) based genotyping results revealed circulation of genotypes of G10 and G3 specificity and P[1] and P[3] types in the region. In humans, only the G1 genotype was detected and for the first time group B rotavirus was detected in an adult. Based on the typical migration pattern of RNA genome segments in the gel electrophoresis (5:2:2:2), group D rotavirus was also identified in poultry diarrheic samples. The results emphasize the need for extended molecular surveys for understanding the interspecies transmission, evolution and especially in designing future vaccine.
INTRODUCTION
Acute diarrhea has been recognized as one of the foremost causes of death in neonates of animal and human globally (Estes and Kapikian, 2007; Dhama et al., 2009). As per the National Animal Health Monitoring System (NAHMS), 2011 report, diarrhea is responsible for more than a half of the mortality (52%) in animals. Rotavirus, the member of family Reoviridae, has a triple layered protein capsid of 75 nm, surrounding the viral genome consisting of 11 segments of double–stranded RNA, which encodes six structural (VP1, VP2, VP3, VP4, VP6 and VP7) and six non–structural proteins (NS1, NS2, NS3, NS4, NS5 and NS6) (Greenberg and Estes, 2009). Rotaviruses have been classified into seven serological species or groups (A–G) based on the antigenic properties of inner capsid protein (VP6) according to the International Committee on Taxonomy of Viruses (ICTV) (Ball et al., 2005). The RVs belonging to species A, B and C (RVA, RVB and RVC, respectively) are known to infect human and various animals, whereas species D, E, F and G (RVD, RVE, RVF and RVG, respectively) thus far have only been recovered from animals, mostly birds (Ball et al., 2005; Matthijnssens et al., 2010). Epidemiologically, RVA is the most important cause of human and animal diarrhea. Specifically, RVA strains have been categorized based on (i) the antigenic properties of VP6, VP7 and VP4 (subgroups, G–serotypes and P–serotypes, respectively); (ii) the migration pattern of the RNA genome segments when subjected to polyacrylamide gel electrophoresis (long, short, super–short or atypical electropherotypes) and (iii) nucleotide sequence analysis (genotypes) (Estes and Kapikian, 2007). To date, 27 G types and 35 P types of RVA have been identified in humans and animals ( Matthijnssens et al., 2011).
The update on prevalence of various genotypes of RVs in different host species belonging to distant geographical locations of India is required for better understanding of interspecies transmission; evolution and in designing vaccine, but several geographical locations in India remain unexplored, with regard to detection and prevalence of RV genotypes. To further explore the epidemiology and genotypic diversity of RV in Madhya Pradesh state, part of central India, we report the findings of RV prevalence and genotypes distribution in different livestock animals and human.
MATERIALS AND METHODS
Clinical Samples and Processing
A total of 733 stool samples were collected from buffaloes (309), cattle (105), goats (25), pigs (69) and poultry (139) in addition to diarrheic human patients (n=86) during April, 2004 to August, 2008 from Jabalpur and surrounding areas of Madhya Pradesh, central India. Majority of these samples were collected from young ones of animals (less than 6 months of age) or children below 5 years of age who were hospitalized with suspected rotavirus gastroenteritis or visited hospital for treatment of acute dehydration due to enteric infection. All the samples were processed by making 10% (w/v) suspension in phosphate buffered saline (pH 7.2) and centrifuged at 10,000g for 30 min to remove course materials. Supernatants were filtered through 0.45 µm syringe filters before storage at –20oC for further use.
Electrophoretic Characterization of dsRNA
The collected stool samples were processed for the detection and confirmation of rotavirus by RNA–PAGE and the genome segments were visualized by silver staining as described in our previous studies (Malik et al., 2011, 2012). The extracted RNA was assessed qualitatively and quantitatively using a Nanodrop Spectrophotometer (ND–1000, ThermoScientific, USA).
Reverse Transcription–polymerase Chain Reaction (RT–PCR)
The first strand cDNA synthesis following PCR for the amplification of full length VP7 (1062 bp) and partial length VP4 gene (864 bp) was carried out as described in our previous studies (Malik et al., 2012; 2013) using the published primers for VP7 (Taniguchi et al., 1992) and VP4 (Isegawa et al., 1993). The multiplex nested PCR was done for genotyping as per the method and primers used in earlier studies (Gouvea et al., 1990, 1994; Isegawa et al., 1993; Iturriza–Gomara et al., 2004). Amplified products were resolved by agarose gel electrophoresis (1% w/v) at 100 V for 1 hour in 1X TAE buffer with 0.5 µg/ml ethidium bromide and viewed under UV trans–illuminator and documented. Amplified products were directly sequenced for VP7 and VP4 genes using the Big dye terminator sequencing kit v3.0 by automated Genetic analyzer ABI 3130xl (Applied Biosystems).
RESULTS AND DISCUSSION
Enteric infections with severe diarrhea are accountable for more than 5% of all deaths among children below 5 years of age and in animals mainly below 6 months of age. Viral pathogens are the most common cause of gastroenteritis in both developing and developed countries (Boschi–Pinto et al., 2008; Dhama et al., 2009; Black et al., 2010) especially rotaviruses (WHO, 2007, 2008; Ramani and Kang, 2007; Tanaka et al., 2007, Parashar et al., 2009; Tate et al., 2009), noroviruses (Radford et al., 2004; Scipioni et al., 2008), astroviruses (De Benedictis et al., 2011) and adenoviruses (Baber and Candy, 1981; McFerran and Smyth, 2000). Rotaviruses belonging to species A, B and C (RVA, RVB and RVC, respectively) are known to infect humans and various animals, whereas species D, E, F and G (RVD, RVE, RVF and RVG, respectively) thus far have only been recovered from animals, mostly birds (Ball et al., 2005; Matthijnssens et al., 2010; Otto et al., 2012). Epidemiologically, RVA is the most important cause for animal as well as human diarrhea.
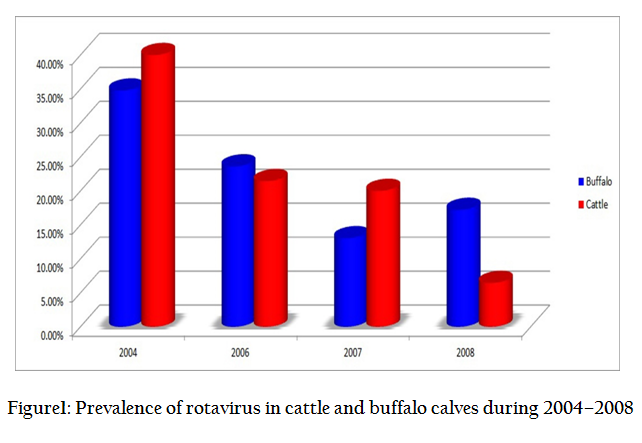
Prevalence of Rotavirus in Cattle and Buffalo Calves
Though extensive epidemiological studies have described the occurrence of RVs from animals in India (Minakshi et al., 2005, 2009; Ghosh et al., 2008; Manuja et al., 2008; Nataraju et al., 2009; Basera et al., 2010; Kusumakar et al., 2010; Dash et al., 2011; Rai et al., 2011; Singh and Jhala, 2011; Malik et al., 2012), still the information on prevalence and genotypes distribution of RV from central part of India is scary. The samples collected during the period of April–June, 2004 revealed the presence of group A rotavirus in 34.78% (24/69) of diarrheic buffalo calves, while 2 out of 5 diarrheic samples from cattle calves were also found positive for RV based on genomic pattern migration in RNA–PAGE. Age wise, higher susceptibility to RV infection was seen in calves of 3–6 weeks old age group (55.55%) followed by up to 3 weeks of age (36.36%). Sex wise, male calves (42.85%) were found more susceptible compared to female calves (28.2%). Subsequently, in 2006 sampling was done to know the prevalence of RVs in animals, in which maximum prevalence was recorded in buffalo calves (23.61%, 17/72) than cattle calves (21.43%, 6/28). In both the study periods, younger ones of less than 3 months of age were more susceptible to virus infection. In contrast to previous year report, in which male calves showed higher susceptibility to infection, during 2006, female calves (25.32%) were slightly more susceptibility than males (22.39%). In 2007, 13.3% (6/46) buffalo calves were found positive in comparison to 20% (2/10) cattle calves. In 2008, the prevalence of RV in buffalo and cattle calves was 17.2% (21/122) and 6.45% (4/62), respectively. Overall, RV prevalence was 22.01 % in buffaloes and 13.33% in cattle calves over the period, 2004–2008 (Figure 1).
Prevalence of Rotavirus in Piglets
In 2006, sampling was done from diarrheic piglets to know the prevalence of RV in swine population of the state. Group A rotavirus was detected in 25.7% piglets (9/35), where all RV positive samples showed typical migration pattern of 4:2:3:2 in RNA–PAGE. Surprisingly, in the subsequent year (2008) all the diarrheic piglets (n=34) were found negative for RV infection, which need further evaluation.
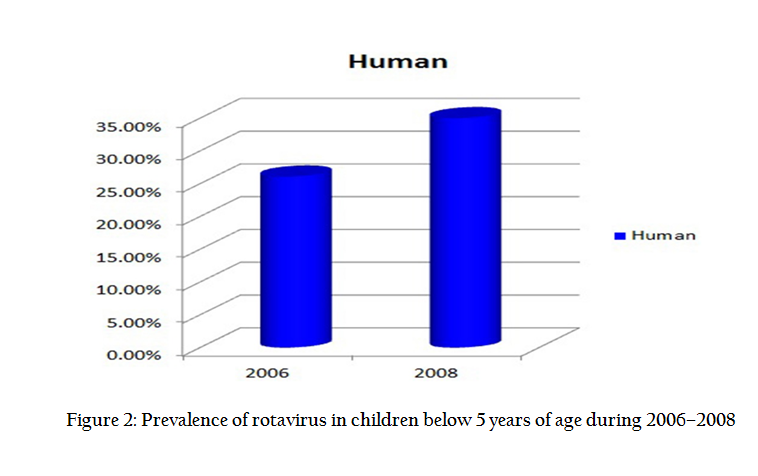
Prevalence of Rotavirus in Human
In 2006, sampling was done to know the prevalence of RVs in animals as well as humans and its cross species transmission possibility, in which 26.09% (12/46) prevalence of RV was recorded in diarrheic children. Younger ones of less than 1 year of age were found more susceptible to RV infection. In 2008, 35% (14/40) of samples from children were detected positive. All the RV positive samples exhibited 4:2:3:2 type migration pattern (group A rotavirus), while one sample of adult human showed 4:3:3:1 indicative of group B rotavirus. Also long electropherotypes were observed in all expect two human samples in which short electropherotypes were observed. Overall, RV prevalence in human was 30.23% over the period, 2006–2008 (Figure 2).
Prevalence of Rotavirus in Poultry
The study on avian RV prevalence was conducted during 2006 and virus was detected in 11.68% (9/77) of faecal samples based on migration pattern in RNA–PAGE. More than 75% of birds below 3 months of age showed RV infection, and broilers were comparatively more susceptible to infection than layers. Interestingly, RNA–PAGE profiles of genomic RNA segments of avian RVs revealed the presence of two different groups i.e. group A (5:1:3:2) in seven samples and group D rotaviruses (5:2:2:2) in two samples. This was the first report of group D rotavirus from poultry in India based on genomic profile in PAGE after silver staining (Savita et al., 2008).
Overall, the prevalence of RVs in central India (Figure 3a,b) was found maximum in human (30.23%, 26/86), followed by buffaloes (22.01%, 68/309), cattle (13.33%, 14/105), pigs (13.04%, 9/69) and poultry (6.47%, 9/139).
_138.PNG)
_138.PNG)
Figure 3(b): Percent of India’s population residing in Madhya Pradesh with prevalence of rotavirus in different species of animals and human
In past, it has been observed that the under–five child mortality rate is highest in Madhya Pradesh (89 deaths/1000 live births), approximately three times higher than the Indian under–five child mortality rate (30.15 deaths/1000 live births) (Sample Registration System, Office of Registrar General, India, 7th July, 2011) which could be associated with the higher prevalence of rotavirus like fatal infections in children of the state. Though diarrheic samples collected from goats were found negative for RV, but needs further investigation on large number of sample size for the exact scenario of its existence from the region.
Genotypic Diversity
The rapid evolution of RVs by means of various mechanisms especially through the exchange of genomic segments among the different host specific RVs lead to diverse genotypes and emergence of novel RVs strains having the capability to jump cross–species barrier (Jagannath et al., 2000; Laird et al., 2003; Esona et al., 2004, 2009, 2010; Mascarenhas et al., 2007; Matthijnssens et al., 2008, 2009; De Donno et al., 2009; Bonkoungou et al., 2011; Mukherjee et al., 2012). In India, during late 1990’s, the first report on animal group A rotavirus genotypes distribution was published by Gulati et al. (1999) based on G & P typing of rotavirus–positive samples collected during 1994 and 1997, in which G10 was found predominating (83%) over others G6 (6%). The majority (94%) of bovine RVs had P[11] while one isolate exhibited P[1] genotype specificity. The most common combination of G and P types detected was G10P[11] (81%), followed by G6P[1] (3%) and G6P[11] (3%). Since, the first report on genotypic distribution of bovine RVs in 1997, it has been identified in diarrheic calves from northern region: Kashmir (G6, G8 and G10), G10P[11] G8+G10P[11]; G10P[11], Haryana and adjoining region (Minakshi et al., 2005; Malik et al., 2013) with predominance of G10 genotype (Basera et al., 2010; Rai et al., 2011), G6P[11] and G10P[3]), Haryana and Punjab (Manuja et al., 2008); Eastern region: G3P[3] strains from Kolkata (Ghosh et al., 2007) [G10]; Western region: G8P[14] strains from Pune (Chitambar et al., 2011). Novel and/or unusual bovine group A RV strains have also been reported viz. G10P[3] from northern (Manuja et al., 2008) and G8P[14] from western India (Chitambar et al., 2011). Rare G15P[11] and G15P[21] strains have been identified and characterized from eastern India (Ghosh et al., 2008). G10P[11] has been found in asymptomatic and symptomatic infections in Indian children, and thought to be acquired through zoonotic transmission (Rao et al., 2000; Varshney et al., 2002; Iturriza–Gomara et al., 2004).
During the study period, genomic diversity was studied based on both genomic RNA migration pattern in RNA–PAGE and multiplex nested PCR. The RV isolates detected during 2006 revealed eight different electropherotypic patterns in animals i.e. four in buffaloes, three in cattle, one in swine and five electropherotypic patterns in human. Interestingly, two unique electropherotypes (one common in cattle and swine; another common in buffalo, cattle and humans) were found suggestive of possible cross–species transmission of RVs. Subsequently, in 2007, all (n=8) the RNA–PAGE positive RV samples were genotyped as of G10 specificity, while none of the sample was typed in nested RT–PCR assay for P–genotype. Studies conducted with bovine RV recovered worldwide showed that serotype G6 and G10 predominate in cattle, although samples with specificity to serotypes G1, G3, can also be found (Suresh et al., 2012). The most frequent RV P genotypes detected in bovine faeces are P[1]; P[5] and P[11] (Gulati et al., 1999; Rao et al., 2000; Minakshi et al., 2005; Ghosh et al., 2007; Manuja et al., 2008; Chitambar et al., 2011; Malik et al., 2012; Suresh et al., 2012). Differences were more frequent in the migration of 10th and 11th segments of RV isolates from cattle, buffalo and human. Both short and long electropherotypes were recorded. In human, the most common genotypes identified are G1 genotype. More recently,
Malik et al. (2012) described the genotypic distribution of RVs in Indian bovine population with unexpectedly higher proportions of G3 genotype. PCR–genotyping confirmed that 39.4% (13/33) of the prevalent RVs were the G3 type and P typing revealed that 93.9% (31/33) of the samples were P[11]. The changing pattern of genotypic circulation in bovines over the time further needs more extensive molecular epidemiological studies in other species also so as to reach a stage where precise situation of most predominant genotypes can be known. In this study, we found the common bovine rotavirus of G10 genotype in central India and in humans it was of G1 specificity.
CONCLUSIONS
The present findings conclude that the occurrence of rotavirus in bovine, porcine, poultry and human population in central India is quite consistent. It also adds to the epidemiological data on rotavirus genotypes circulating in dairy herds of Madhya Pradesh, central India. However, still more elaborate studies should be conducted to estimate the gravity of the situation as well as the epidemiological understanding of contributing viral genotypes in future for the development and implementation of efficient immunization approaches, thereby controlling infection and reducing economic losses.
ACKNOWLEDGMENTS
The authors wish to acknowledge the help received from State Animal Husbandry Department during collection of samples and MP Rural Livelihood Project and MP Council of Science and Technology for financial help.
REFERENCES
Baber DJ and Candy JB (1981). Isolation and characterization of bovine adenoviruses types 3, 4 and 8 from free living African buffaloes (Syncerus caffer). Res. Vet. Sci. 31(1): 69–75.
PMid:7313323
Ball JM, Mitchell DM, Gibbons TF and Parr RD (2005). Rotavirus NSP4: a multifunctional viral enterotoxin. Viral Immunol. 18: 27–40.
http://dx.doi.org/10.1089/vim.2005.18.27
PMid:15802952
Basera SS, Singh R, Vaid N, Sharma K, Chakravarti S and Malik YPS (2010). Detection of Rotavirus Infection in Bovine Calves by RNA–PAGE and RT–PCR. Ind. J. Virol. 21(2):144–147.
http://dx.doi.org/10.1007/s13337-010-0017-9
PMid:23637494 PMCid:PMC3550708
Black RE, Cousens S, Johnson HL, Lawn JE, Rudan I, Bassani DG, Jha P, Campbell H, Walker CF, Cibulskis R, Eisele T, Liu L, Mathers C; Child Health Epidemiology Reference Group of WHO and UNICEF (2010). Global, regional, and national causes of child mortality in 2008: a systematic analysis. Lancet 375: 1969–1987.
http://dx.doi.org/10.1016/S0140-6736(10)60549-1
Bonkoungou IJ, Damanka S, Sanou I, Tiendrébéogo F, Coulibaly SO, Bon F, Haukka K, Traoré AS, Barro N and Armah GE (2011). Genotype Diversity of Group A Rotavirus Strains in Children With Acute Diarrhea in Urban Burkina Faso, 2008–2010. J. Med. Virol. 83: 1485–1490.
http://dx.doi.org/10.1002/jmv.22137
PMid:21678452
Boschi–Pinto C, Velebit L and Shibuya K (2008). Estimating child mortality due to diarrhoea in developing countries. Bull. World Health Org. 86(9): 710–717.
http://dx.doi.org/10.2471/BLT.07.050054
PMid:18797647 PMCid:PMC2649491
Chitambar SD, Arora R, Kolpe AB, Yadav MM and Raut CG (2011). Molecular characterization of unusual bovine group A rotavirus G8P[14] strains identified in western India: Emergence of P[14] genotype. Vet. Microbiol. 148: 384–388.
http://dx.doi.org/10.1016/j.vetmic.2010.08.027
PMid:20880637
Dash SK, Tewari A, Kumar K, Goel A and Bhatia AK (2011). Detection of Rotavirus from diarrhoeic cow calves in Mathura, India. Vet. World 4(12): 554–556.
http://dx.doi.org/10.5455/vetworld.2011.554-556
De Benedictis P, Schultz–Cherry S, Burnham A and Cattoli G (2011). Astrovirus infections in humans and animals – molecular biology, genetic diversity, and interspecies transmissions. Infect. Genet. Evol. 11(7): 1529–1544.
http://dx.doi.org/10.1016/j.meegid.2011.07.024
PMid:21843659
De Donno A, Grassi T, Bagordo F, Idolo A, Cavallaro A, Gabutti G and Collaborative Group for the surveillance of Rotavirus Infection (2009).Emergence of unusual human rotavirus strains in Salento, Italy, during 2006–2007. BMC Infect. Dis. 9: 43.
http://dx.doi.org/10.1186/1471-2334-9-43
PMid:19368717 PMCid:PMC2676288
Dhama K, Chauhan RS, Mahendran M and Malik SVS (2009). Rotavirus diarrhea in bovines and other domesticated animals. Vet. Res. Comm. 33(1): 1–23.
http://dx.doi.org/10.1007/s11259-008-9070-x
PMid:18622713
Esona MD, Armah GE, Geyer A and Steele AD (2004). Detection of an Unusual Human Rotavirus Strain with G5P[8]Specificity in a Cameroonian Child with Diarrhea. J. Clin. Microbiol. 42(1): 441–444.
http://dx.doi.org/10.1128/JCM.42.1.441-444.2004
PMid:14715801 PMCid:PMC321728
Esona MD, Geyer A, Banyai K, Page N, Aminu M, Armah GE, Hull J, Steele DA, Glass RI and Gentsch JR (2009). Novel Human Rotavirus Genotype G5P[7] from Child with Diarrhea, Cameroon. Infect. Genet. Evol. 15(1): 83–86.
Esona MD, Page NA, Akran VA, Armah GE and Steele AD (2010). Characterization of 2 Human Genotype G10Rotavirus Strains, 3008CM and 1784/CI/1999, Isolated in Cameroon and Cote d'Ivoire during the 1999–2000 Rotavirus Season. J. Infect. Dis. 202(S1): S212–S219.
http://dx.doi.org/10.1086/653553
PMid:20684705
Estes M and Kapikian A (2007). Rotaviruses. In: Knipe DM, Howley PM, Griffin DE, Lamb RA, Martin MA, Roizman B, Straus SE, editors. Fields virology. Philadelphia: Kluwer Health/Lippincott, Williams and Wilkins. p. 1917–1974.
Ghosh S, Samajdar S, Sinha M, Kobayashi N, Taniguchi K and Naik TN (2008). Molecular characterization of rare bovine group A rotavirus G15P[11] and G15P[21] strains from eastern India: identification of simian SA11–like VP6 genes in G15P[21] strains. Virus Genes 37: 241–249.
http://dx.doi.org/10.1007/s11262-008-0260-y
PMid:18679786
Ghosh S, Varghese V, Samajdar S, Sinha M, Kobayashi N and Naik TN (2007). Molecular characterization of bovine group A rotavirus G3P[3] strains. Arch. Virol. 152: 1935–1940.
http://dx.doi.org/10.1007/s00705-007-1009-y
PMid:17577612
Gouvea V, Glass RI, Woods P, Taniguchi K, Clark HF, Forrester B and Fang ZY (1990). Polymerase chain reaction amplification and typing of rotavirus nucleic acid from stool specimens. J. Clin. Microbiol. 28: 276–282.
PMid:2155916 PMCid:PMC269590
Gouvea V, Santos N and Timenetsky MDC (1994). Identification of bovine and porcine rotavirus G types by PCR. J. Clin. Microbiol. 32: 1338– 1340.
PMid:8051263 PMCid:PMC263690
Greenberg HB and Estes MK (2009). Rotaviruses: from pathogenesis to vaccination. Gastroenterology 136(6): 1939–1951.
http://dx.doi.org/10.1053/j.gastro.2009.02.076
PMid:19457420 PMCid:PMC3690811
Gulati BR, Nakagomi O, Koshimura Y, Nakagomi T and Pandey R (1999). Relative frequencies of g and p types among rotaviruses from Indian diarrheic cow and buffalo calves. J. Clin. Microbiol. 37(6): 2074–2076.
PMid:10325385 PMCid:PMC85038
Isegawa Y, Nakagomi O, Nakagomi T, Ishida S, Uesugi S and Ueda S (1993). Determination of bovine rotavirus G and P serotypes by polymerase chain reaction. Mol. Cell. Prob. 7: 277–84.
http://dx.doi.org/10.1006/mcpr.1993.1041
PMid:8232344
Iturriza–Gomara M, Kang G and Gray J (2004). Rotavirus genotyping: Keeping up with an evolving population of human rotaviruses. J. Clin. Virol. 31: 259–265
http://dx.doi.org/10.1016/j.jcv.2004.04.009
PMid:15494266
Jagannath MR, Vethanayagam RR, Reddy BS, Raman S and Rao CD (2000). Characterization of human symptomatic rotavirus isolates MP409 and MP480 having 'long' RNA electropherotype and subgroup I specificity, highly related to the P6[1],G8 type bovine rotavirus A5, from Mysore, India. Arch. Virol. 145: 1339–1357.
http://dx.doi.org/10.1007/s007050070094
PMid:10963341
Kusumakar AL, Savita, Malik YPS, Minakshi and Prasad G (2010). Genomic diversity among group A rotaviruses from diarrheic children, piglets, buffalo and cow calves of Madhya Pradesh. Ind. J. Microbiol. 50(1): 83–88.
Laird AR, Ibarra V, Ruiz–Palacios G, Guerrero ML, Glass RI and Gentsch JR (2003). Unexpected Detection of Animal VP7 Genes among Common Rotavirus Strains Isolated from Children in Mexico. J. Clin. Microbiol. 41(9): 4400–4403.
http://dx.doi.org/10.1128/JCM.41.9.4400-4403.2003
PMid:12958276 PMCid:PMC193830
Malik YPS, Naveen Kumar, Kuldeep Sharma, AA Haq, Amit Kumar and Minakshi Prasad (2013). Sequence and phylogenetic analysis of bovine rotavirus isolates (G6 genotypes) from India. Adv. Vet. Anim. Sci. 1(1): 41–43.
Malik YPS, Sharma K, Vaid N, Chakravarti S, Chandrashekar KM, Basera SS, Singh R, Minakshi, Prasad S, Gulati BR, Bhilegaonkar KN and Pandey AB (2012). Frequency of group A rotavirus with mixed G and P genotypes in bovines: predominance of G3 genotype and its emergence in combination with G8/G10 types. J. Vet. Sci. 13(3): 271–278.
http://dx.doi.org/10.4142/jvs.2012.13.3.271
PMid:23006956 PMCid:PMC3467402
Malik YS, Chandrashekar KM, Sharma K, Haq AA, Vaid N, Chakravarti S, Batra M, Singh R and Pandey AB (2011). Picobirnavirus detection in bovine and buffalo calves from foothills of Himalaya and Central India. Trop. Anim. Health Product. 43(8): 1475–1478.
http://dx.doi.org/10.1007/s11250-011-9834-0
PMid:21479844
Manuja BK, Prasad M, Manuja A, Gulati BR and Prasad G (2008). A novel genomic constellation (G10P[3]) of group A rotavirus detected from buffalo calves in northern India. Virus Res. 138: 36–42.
http://dx.doi.org/10.1016/j.virusres.2008.08.006
PMid:18793686
Mascarenhas JD, Leite JP, Lima JC, Heinemann MB, Oliveira DS, Araújo IT, Soares LS, Gusmão RH, Gabbay YB and Linhares AC (2007). Detection of a neonatal human rotavirus strain with VP4 and NSP4 genes of porcine origin. J. Med. Microbiol. 56(4): 524–32
http://dx.doi.org/10.1099/jmm.0.46635-0
PMid:17374895
Matthijnssens J, Ciarlet M, McDonald SM, Attoui H, Bányai K, Brister JR, Buesa J, Esona MD, Estes MK, Gentsch JR, Iturriza–Gómara M, Johne R, Kirkwood CD, Martella V, Mertens PP, Nakagomi O, Parre-o V, Rahman M, Ruggeri FM, Saif LJ, Santos N, Steyer A, Taniguchi K, Patton JT, Desselberger U and Van Ranst M (2011).Uniformity of rotavirus strain nomenclature proposed by the Rotavirus Classification Working Group (RCWG). Arch. Virol. 156(8): 1397–1413.
http://dx.doi.org/10.1007/s00705-011-1006-z
PMid:21597953 PMCid:PMC3398998
Matthijnssens J, Heylen E, Zeller M, Rahman M, Lemey P and Van Ranst M (2010). Phylodynamic analyses of rotavirus genotypes G9 and G12 underscore their potential for swift global spread. Mol. Biol. Evolut. 27(10): 2431–2436.
http://dx.doi.org/10.1093/molbev/msq137
PMid:20522727
Matthijnssens J, Potgieter CA, Ciarlet M, Parre-o V, Martella V, Bányai K, Garaicoechea L, Palombo EA, Novo L, Zeller M, Arista S, Gerna G, Rahman M and Van Ranst M (2009). Are Human P[14] Rotavirus Strains the Result of Interspecies Transmissions from Sheep or Other Ungulates That Belong to the Mammalian OrderArtiodactyla? J. Virol. 83(7): 2917–2929.
http://dx.doi.org/10.1128/JVI.02246-08
PMid:19153225 PMCid:PMC2655590
Matthijnssens J, Rahman M and Van Ranst M (2008). Two out of the 11 genes of an unusual human G6P[6] rotavirus isolate are of bovine origin. J. Gen. Virol. 89: 2630–2635.
http://dx.doi.org/10.1099/vir.0.2008/003780-0
PMid:18796733
McFerran JB and Smyth JA (2000). Avian adenoviruses. Rev. Sci. et Tech. Inter. Off. Epiz. 19(2): 589–601
Minakshi, Prasad G and Grover YP (2009). Occurrence of dual infection of bovine group A rotavirus in diarrhoeic calf in Haryana, India. Ind. J. Anim. Sci. 79 (12): 1205–1208.
Minakshi, Prasad G, Malik Y and Pandey R (2005). G and P genotyping of bovine group A rotaviruses in faecal samples of diarrhoeic calves by DIG–labelled probes. Ind. J. Biotechnol. 4: 93–99.
Mukherjee A, Nayak MK, Roy T, Ghosh S, Naik TN, Kobayashi N and Chawla–Sarkar M (2012). Detection of human G10 rotavirus strains with similarity to bovine and bovine–like equine strains from untypable samples. Infect. Genet. Evol. 12: 467–470.
http://dx.doi.org/10.1016/j.meegid.2011.11.007
PMid:22155585
Nataraju SM, Chattopadhyay UK and Krishnan T (2009). A study on the possibility of zoonotic infection in rotaviral diarrhoea among calves and buffalo calves in and around Kolkata, India. Eur. Rev. Med. Pharmacol. Sci. 13: 7–11.
PMid:19364081
Otto PH, Ahmed MU, Hotzel H, Machnowska P, Reetz J, Roth B, Trojnar E and Johne R (2012). Detection of avian rotaviruses of groups A, D, F and G in diseased chickens and turkeys from Europe and Bangladesh. Vet. Microbiol. 156: 8–15.
http://dx.doi.org/10.1016/j.vetmic.2011.10.001
PMid:22079218
Parashar UD, Burton A, Lanata C, Boschi–Pinto C, Shibuya K, Steele D, Birmingham M and Glass RI (2009). Global mortality associated with rotavirus disease among children in 2004. J. Infect. Dis. 200(1): 9–15.
http://dx.doi.org/10.1086/605025
PMid:19817620
Radford AD, Gaskell RM and Hart CA (2004). Human norovirus infection and the lessons from animal caliciviruses. Curr. Opin. Infect. Dis. 17(5): 471–478.
http://dx.doi.org/10.1097/00001432-200410000-00012
PMid:15353967
Rai RB, Hansha A, Rai S, Singh B, Kumar H, Singh AK, Damodaran T and Dhama K (2011). Prevalence of rota and coronavirus infections in calves of Barabanki and Raebareli districts of Uttar Pradesh. Ind. J. Vet. Pathol. 35(1): 73–74.
Ramani S and Kang G (2007). Burden of disease and molecular epidemiology of group A rotavirus infections in India. Ind. J. Med. Res. 125(5): 619–632.
PMid:17642497 PMCid:PMC2474761
Rao CD, Gowda K and Reddy BS (2000). Sequence analysis of VP4 and VP7 genes of nontypeable strains identifies a new pair of outer capsid proteins representing novel P and G genotypes in bovine rotaviruses. Virology 276: 104–113.
http://dx.doi.org/10.1006/viro.2000.0472
PMid:11021999
Savita, Kusumakar AL, Malik YPS, Minakshi and Prasad G. 2008). Detection and characterization of group A and D avian rotaviruses in India. Ind. J. Biotechnol. 7: 554–556.
Scipioni A, Mauroy A, Vinjé J and Thiry E (2008). Animal noroviruses. Vet. J. 178(1): 32–45.
http://dx.doi.org/10.1016/j.tvjl.2007.11.012
PMid:18294883
Singh TC and Jhala MK (2011). G–typing of bovine rotaviruses by using VP7 gene specific heminested RT–PCR from diarrhoeic calf faecal samples. Buff. Bull. 30(2): 113–138.
Suresh T, Rai RB, Dhama K, Sawant PM, Kumar D and Bhatt P (2012). Determination of G and P type diversity of group A rotaviruses and detection of a new genotype from diarrhoeic calves in northern and southern states of India. Vet. Pract. 13(1): 1–8.
Tanaka G, Faruque AS, Luby SP, Malek MA, Glass RI and Parashar UD (2007). Deaths from rotavirus disease in Bangladeshi children: estimates from hospital–based surveillance. Ped. Infect. Dis. J. 26(11): 1014–1018.
http://dx.doi.org/10.1097/INF.0b013e318125721c
PMid:17984808
Taniguchi K, Wakasugi F, Pongsuwanna Y, Urasawa T, Ukae S, Chiba S and Urasawa S (1992). Identification of human and bovine rotavirus serotypes by polymerase chain reaction. Epidemiol. Infect. 109: 303–312.
http://dx.doi.org/10.1017/S0950268800050263
PMid:1327857 PMCid:PMC2271918
Tate JE, Chitambar S, Esposito DH, Sarkar R, Gladstone B, Ramani S, Raghava MV, Sowmyanarayanan TV, Gandhe S, Arora R, Parashar UD and Kang G (2009). Disease and economic burden of rotavirus diarrhoea in India. Vaccine 27(5): 18–24.
http://dx.doi.org/10.1016/j.vaccine.2009.08.098
PMid:19931713
Varshney B, Jagannath MR, Vethanayagam RR, Kodhandharaman S, Jagannath HV, Gowda K, Singh DK and Rao CD (2002). Prevalence of, and antigenic variation in, serotype G10 rotaviruses and detection of serotype G3 strains in diarrheic calves: implications for the origin of G10P11 or P11 type reassortant asymptomatic strains in newborn children in India. Arch. Virol. 147(1): 143–165.
http://dx.doi.org/10.1007/s705-002-8308-z
PMid:11855628
World Health Organization (2007). Rotavirus vaccines: WHO position paper. Weekly Epidemiol. Rec. 82(32): 285–296.
PMid:17691162
World Health Organization (2008). Global networks for surveillance of rotavirus gastroenteritis, 2001–2008. Weekly Epidemiol. Rec. 83(47): 421–428.
PMid:19024780



