Advances in Animal and Veterinary Sciences
Research Article

Histomorphological Study of Syrinx of Black Francolin (Francolinus francolinus) in Iraq
Muna Hussain Al-aameli1, Khalid Kamil Kadhim2*
1Department of Anatomy and Histology, Faculty of Veterinary Medicine, University of Kerbala-Kerbala-Iraq; 2Department of Anatomy and Histology, Faculty of Veterinary Medicine, University of Baghdad-Baghdad-Iraq.
Abstract | The syrinx of male and female adult black francolin was studied. Seven adult males and females weighted (300) gm to (265) gm respectively, were captured from the village of Baghdad. Macroscopic examination of syrinx was done under using stereomicroscope image analysis. Histological sections stained with heamatoxylin and eosin stain, Massson’s trichrom stain; Periodic acid schiff; Alcian blue stain; and combined Aldehyde fuchsin-Alcian blue stain. Results, the syrinx was tracheobronchial in type; it was consisted from: Tympanum, Tracho-Syringeal part, and Brancho-Syringeal part. In addition, there were lateral and medial vibrating membranes, pessulus and triangular voice. The syrinx was lined by the pseudostratified ciliated columnar epithelia with goblet cells. Groups of goblet cells are formed intra epithelia mucous glands with sulphated mucin. Complete cartilaginous rings were formed upper part of syringeal skeleton, whereas, C shaped cartilages formed its middle and distal parts. Medial and lateral labia were protruded into the syringeal lumen. Sexual dimorphism were obtained, the males had significantly narrow tympanum and right syringeal passage with thicker syringeal labia than female.
Keywords | Black francolin, Tympanum, Vibrating membrane, Syrinx, Labia
Editor | Kuldeep Dhama, Indian Veterinary Research Institute, Uttar Pradesh, India.
Received | January 10, 2017; Accepted | February 16, 2017; Published | February 28, 2017
*Correspondence | Khalid Kamil Kadhim, Department of Anatomy and Histology, Faculty of Veterinary Medicine, University of Baghdad-Baghdad-Iraq; Email: khalidkamkad@yahoo.com
Citation | Al-Aameli MH, Kadhim KK (2017). Histomorphological study of syrinx of black francolin (francolinus francolinus) in Iraq. Adv. Anim. Vet. Sci. 5(2): 92-99.
DOI | http://dx.doi.org/10.14737/journal.aavs/2017/5.2.92.99
ISSN (Online) | 2307-8316; ISSN (Print) | 2309-3331
Copyright © 2017 Al-Aameli and Kadhim. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Introduction
Birds have evolved a unique phonatory organ (syrinx), which is highly variable in structure between species. Like the mammalian larynx, regulates airflow and action influences acoustic parameters of syrinx is controlled by muscles during closing and opening of the passage airways (Andersson, 1994; Podos et al., 2004; Edwards et al., 2005). The classification of birds depended on presence or absence the basis structure of syrinx or their musculature (Tsukahara et al., 2008). Some birds vocalize all year long while others call only during the mating season (Zhang et al., 2009). And this is also accompanied by alteration in the functional morphology of certain organs that responsible for sound generation. Nevertheless, to what level of gender dimorphism that affects the control vocal center, it is still indistinct. Tympanum differs in birds, its consists from two rings in the Japanese quails (Cevik-Demirkan et al., 2007) and five in the pigeons (Yildiz et al., 2005). Histological and morphological syringeal variation in the songbird has not been inspected. However, such changes have been confirmed in mammalian species where they lead to clear gender dimorphism in the characters of voice (Suthers, 2004). In the songbirds the labia play a major role in sound generators. Particularly, laterally location labia are effective into vibration (Riede and Goller, 2010). The wedge-shaped pessulus divided the airways of syrinx (Kabak et al., 2007). The males black francolin have in addition to its exterior differences with female, they have characteristic high loud voice especially in breeding season unlike females that keep with inaudible sound (Prince et al., 2011). There is a dearth of information concerning the anatomical and histological aspect of syrinx in this bird, and whether there is any difference between the sexes, especially that males have characteristic high voice. For this reason, this experiment was designed.
Materials and methods
Seven males and females of black francolin weighted (300) gm to (265) gm respectively, which captured from the village of capital Baghdad were used in this study. The birds euthanized by intra muscular administration of diazepam (1mg/1kg) combined with ketamine HCL (30Mg, 1Kg). After thoracic incision, the syrinx are collected and fixed in the 10% neutral buffer formalin. Anatomical examination was done using dissection microscope (Kruss Ootronic, Germany) provided by camera (SMZ 1500 digital camera). Longitudinal and cross sections were prepared histologically. Routine heamatoxylin and eosin stain were used in addition to other special stain (Massson’s trichrom stain; PAS; Alcian blue stain; Combined Aldehyde fuchsin-Alcian blue method) to give more histological details, and Alcian blue-Alizarin red technique to recognize the ossification of hyaline cartilage (Bancroft and Stevens, 2012). The parameters that have been measured are the diameters of syringeal passages and diameters of the lateral trachealis and sternotrachealis. In addition estimation of the thickness of tympanic (vibrating) membranes, labia thickness, pessulus and triangular voice area were done.
Statistical analysis
The collected data were represented by mean ± standard deviation. Comparisons were done between male and female. The mean of differences between data were estimated by independent t test using SPSS version 20.
Result and disscusion
Syrinx of black francolin was tracheobronchial in type; it consists from three different groups: Tympanum, Tracho-Syringeal part, Brancho-Syringeal part (Figure 1 and 2). In addition, there were branchidesmus (interbronchial ligament), lateral and medial vibrating membranes, pessulus and triangular voice. Similar to this syringeal division was mentioned in turkeys (Khaksar et al., 2012). Meanwhile, in the other song birds, the syrinx is consisted from either the tracheal or bronchial part or both (Seller, 2002).
Three incomplete rings were formed tympanum of francolin syrinx. The last rings of tympanum bifurcate into two modified rings to form the tracho-Syringeal part (three rings), this followed by last part of syrinx (Brancho-Syringeal) which consisted from four incomplete rings (Figure 3). Comparing with sea gulls, the tympanum is consisted from five rings (İnce et al., 2012), and in long-legged buzzard are two rings (Kabak et al., 2007).

Figure 1: Macrograph of black francolin syrinx (ventral view): showing the parts of syrinx that formed by: tympanum (tp); trachea-syringeal part (ts); broncho-Syringeal part (bs); triangular voice (tv); ligament (ig); sternotrachilis (st); lateral trachealis (lt); primary bronchi (br).
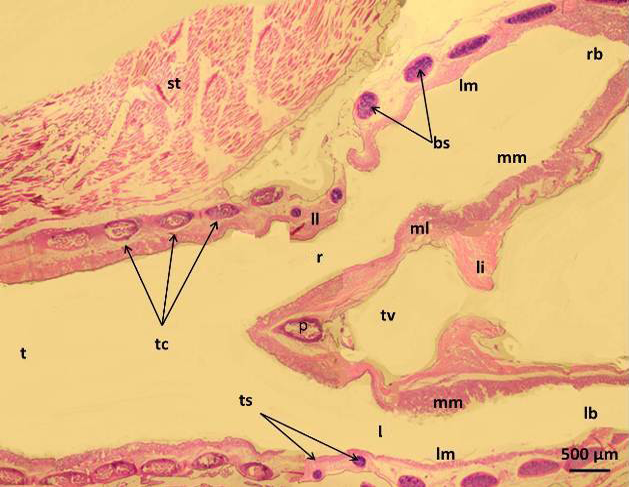
Figure 2: Micrograph of syrinx of male black francolin showing: trachea (t); tympanum cartilages (tc); trachea-syringeal cartilages (ts); pessulus (p); right passage (r); left passage (l); lateral labium (ll); medial labium (ml); lateral vibrating membrane (lm); medial vibrating membrane (mm); triangular voice (tv); sternotrachealis (st): ligament (li). H&E.
In francolin, the tympanum was lined by the pseudostratified ciliated columnar epithelia with goblet cells, the later however tended to aggregates in middle and lower parts of syrinx to form intra epithelia mucous glands which appeared in different numbers and sizes (Figure 4). These glands or the goblet cells showed positive reaction with PAS stain and Alcian blue stain (Figure 5). However, these glands appeared magenta due to its contains of sulphated mucin (Figure 6). In Bursa roller pigeon this mucosa is compose of pseudostratified prismatic epithelium or bistratified squamous, columnar (Yıldız et al., 2005; Demir kan et al., 2006) had recorded that the gland and goblet cells increased toward the broncheo-syringeal part in bursa roller pigeon. In birds, (Bacha and Bacha, 2000) and goose, (Jeffery, 2004) mentioned that plentiful goblet cells constituted the intraepithelial glands which are contain mucin with a predominance of sulphate ester salt for protection.
In the current study, the lamina propria-submucosa was consisted from loose connective tissue with high amount of elastic fibers which showed continuity with the perichondrium that surround the hyaline cartilage. The chondrocytes of the later located inside the lacunae and surrounded by homogenous matrix. These hyaline cartilages were mostly ossified at its center (Figure 7). Similar results were reported by (Aughey and Frye, 2001) however, there was partly ossified perichondrium of the syringeal hyaline cartilages in avian.
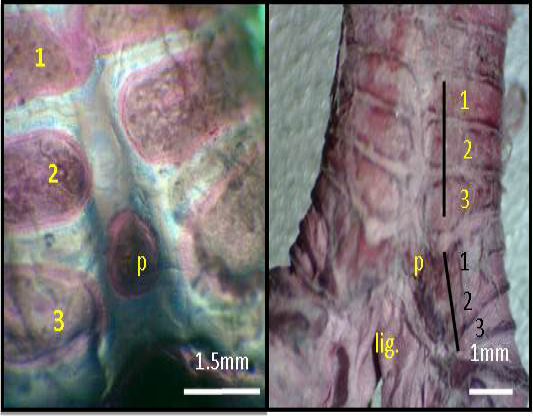
Figure 3: Macrograph of syrinx (dorsal view) of black francolin: showing tympanum which formed by (1, 2, 3, yellow color) C shaped cartilages; trachea-syringeal part which formed by (1, 2, 3, black color) C shaped cartilages; pessulus (p); ligament (lig.) Alcian blue-Alizarin red technique.
The tympanum diameter showed no difference between genders (Figure 8). Meanwhile, the diameter of right passage appeared significantly (P<0.05) narrower in male (Figure 9). In contrast, the left passage showed no difference between genders (Figure 10). These data conceded with (Suthers and Zollinger, 2004) report that in songbird, the left passage is source sound produces lower than the right side.
In the present study, the medial and lateral labia were thickening of the mucous membrane that protruded into the syringeal lumen between tympanum and the broncheo-syringeal parts of syrinx. The medial labia attached with the pesselus cranially and continue with thin medial tympanic (vibrating) membranes caudally, in which the later has attached with the bronchial wall medially. Meanwhile, the syringeal muscles attached directly with medial labia (Figure 11).
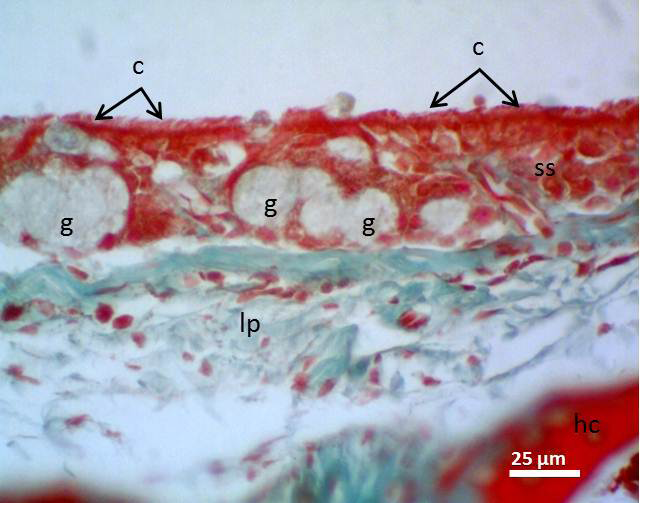
Figure 4: Micrograph of syrinx epithelium of black francolin. Whereas, cilia (c); pseudostratified ciliated columnar epithelia (ss); aggregation of goblet cells (intra epithelia mucous glands) (g); lamina propria (lp); hyaline cartilage (hc). Massson’s trichrom stain.
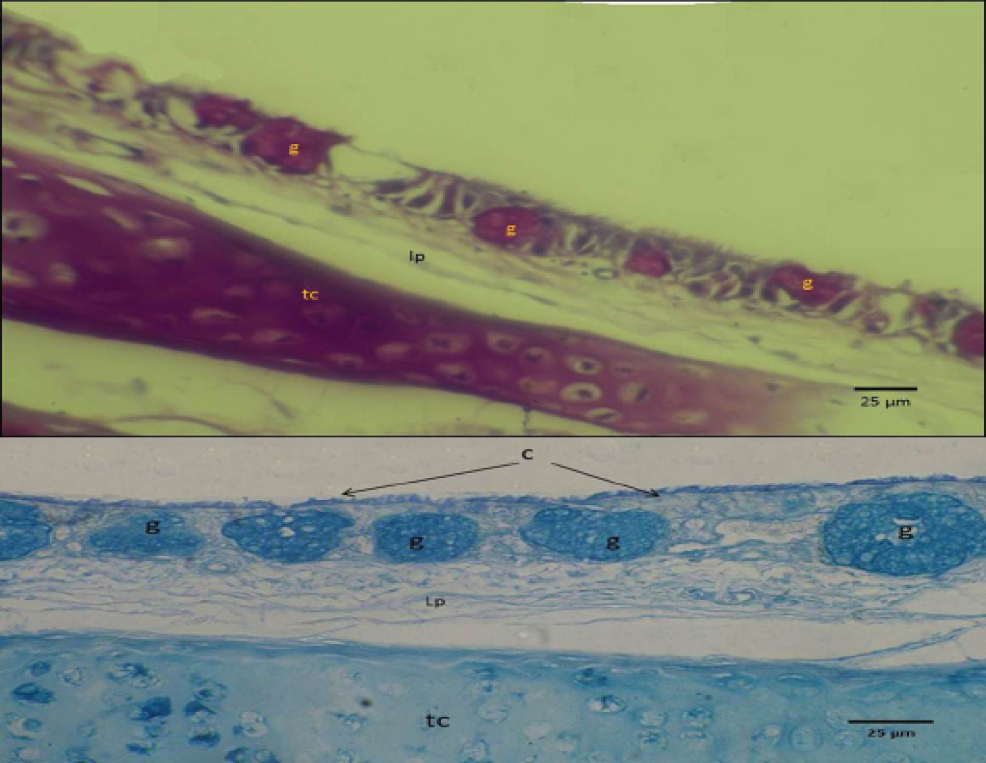
Figure 5: Micrograph of syrinx epithelium of black francolin. Showing mucin reaction of the mucous epithelia (upper with PAS) and (lower with Alcian blue stan), whearas, trachea-syringeal part (ts); aggregation of goblet cells (intra epithelia mucous glands) (g); lamina propria (lp).
The epithelium lining these labia had characteristic features, it not appeared as typical stratified columnar cells, and these cells appeared lower in height and more compact. This may be due to continues direct effects of the air flow on this part of mucosa. The extra cellular matrix of these labia was consisted mainly from collagen and smooth muscle fibers. These fibers were oriented randomly particularly in the superficial layers (Figure 12).
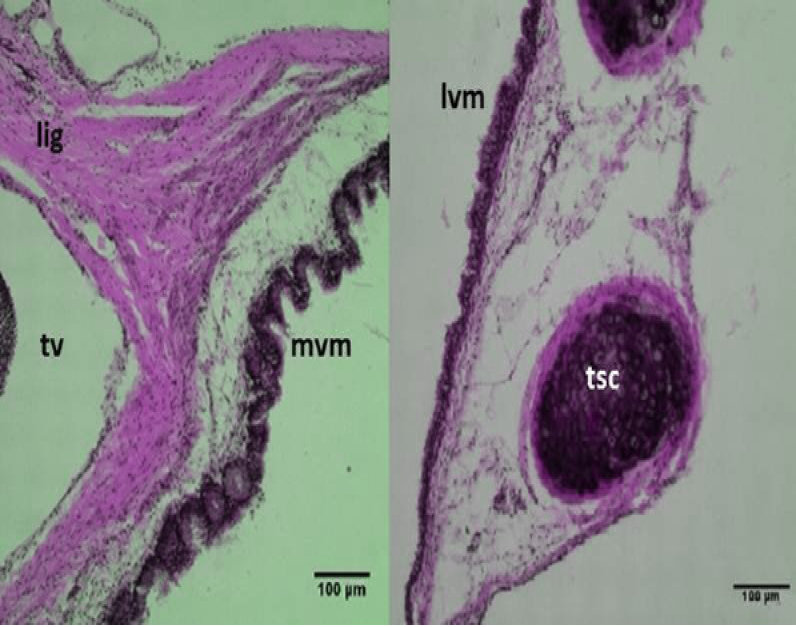
Figure 6 Micrograph: Medial vibrating membranes (mvm) notice the folded; lateral vibrating membranes (lvm); tracheo-syringeal cartilages (tsc); triangular voice (tv); sternotrachealis ligament (lig). (Combined Aldehyde Fuchsin-Alcian blue method).
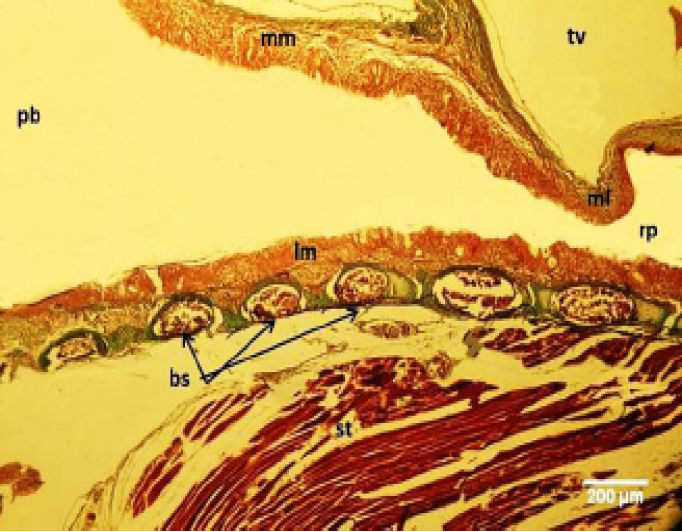
Figure 7: Micrograph of male black francolin syrinx. Showing ossification of the syringeal cartilages (bc); pseudostratified ciliated columnar epithelia lining the lateral vibrating membrane (lm) and medial vibrating membrane (mm); right syringeal passage (rp); primary bronchi (pb); triangular voice (tv); sternotrachealis (st). Massson’s trichrom stain.
.
The right and left medial labia and right lateral labia of male were significantly (P<0.05) thicker than female (Table 1). (Riede and Goller, 2015) recorded that the labium is differ in shape and size between the female and male which is generally thicker in male. Related to these results, (Fee, 2002) suggested that in songbird, the labia of right side is source of sound and has higher resonance peaks
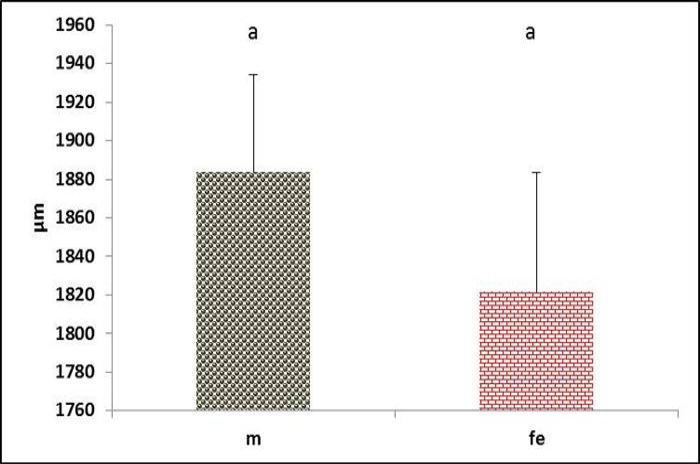
Figure 8 Tympanum diameter in both male and female black francolin. Values represent means±SD. Different letters means significantly (P<0.05) different. Where, male (m); female (fe).
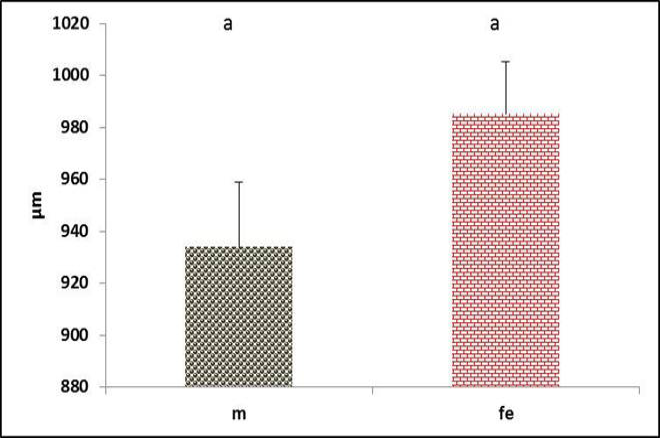
Figure 9: Diameter of the right syringeal passage in both male and female black francolin. Values represent means±SD. Different letters means significantly (P<0.05) different. Where, male (m); female (fe).

Figure 10: Diameter of the left syringeal passage in both male and female black francolin. Values represent means±SD. Different letters means significantly (P<0.05) different. Where, male (m); female (fe).
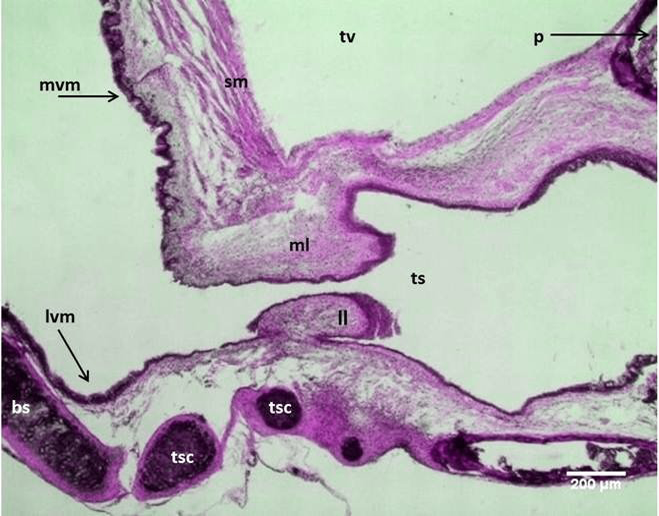
Figure 11: Micrograph: Longitudinal section of the syrinx in the male black francolin showing: medial labia (ml); lateral labia (ll); lateral vibrating membranes (lvm); medial vibrating membranes (mvm); tracheo-syringeal part (ts); tracheo-syringeal cartilages (tsc); triangular voice (tv); syringeal muscle (sm); broncheo-syringeal parts (bs). (Combined Aldehyde Fuchsin-Alcian blue method).
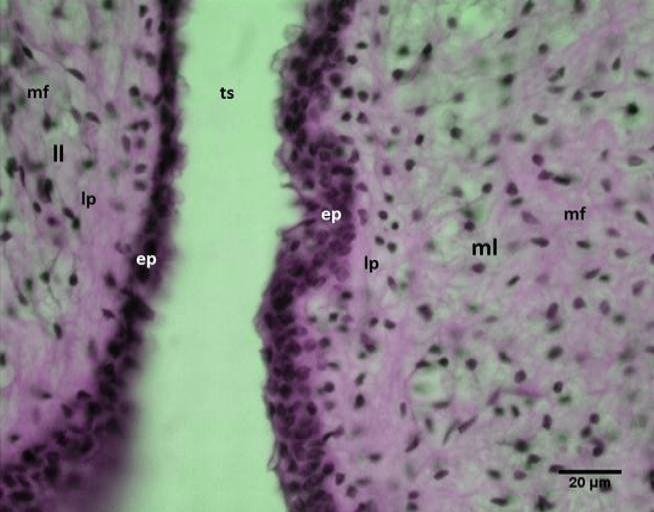
Figure 12: Micrograph: Syringeal labia of the male black francolin showing: medial labia (ml); lateral labia (ll); epithelium (ep); lamina propria (lp); muscle fibers (mf); passage of the tracheo-syringeal part (ts). (Combined Aldehyde Fuchsin-Alcian blue method).
than those of left side. Furthermore, (Gasser et al., 2006) explained that the extra cellular matrix components (elastic and collagen fibers) of this labium were general feature design in organs to accommodate stress. In addition, (Suthers, 2004) recorded that the labia of the syrinx in passerine species appear as cushions of loose connective tissue.
In the current study, the bronchosyringeal part of black francolin syrinx is consisted from four C shaped cartilages. Whereas this part is composed from 7 cartilage rings in se-
Table 1: the thickness of syringeal labia of black francolinus (male and female).
|
Thickness left lateral labia (µm) |
Thickness right lateral labia (µm) |
Thickness left medial labia (µm) |
Thickness right medial labia (µm) |
|
|
Male |
170±6 a |
191±26 a |
176±19 b |
193±10 a |
|
Female |
152±21 a |
156±18 b |
109±22 c |
166±14 b |
The data represented as (mean ± SD), (different letters) mean significant different at (P ≤ 0.05) within rows and between male and female.
a gulls (İnce et al., 2012). In present study, the medial (internal) and lateral (external) vibrating membranes of the brancheosyringeal part were lined by similar epithelia. However, the medial vibrating membrane seemed more folded than the lateral one, may be due to effects of muscle contraction at this part when the hyaline cartilage had replaced medially by thin layers of smooth muscles fibers with fine collagen and elastic fibers (Figures 6). The epithelia of internal and external tympanic membranes in male had dominantly thicker (P<0.05) than female (Figure 13 and 14). Concerning this result, (Tshifhiwa et al., 2011) had suggested that these tympaniforum membrane lined with a bands of connective tissue that varies in thickness depended on species. And this connective tissue could modify the syringeal lumen (Brown and Ward, 1990).
The syrinx had clear skeletal muscles fibers which represented by lateral trachealis and sternotrachealis at each side of the tympanum rings (Figure 1). The diameter of these muscles through cross sections in males were significantly (P˂0.05) higher than females (Figure 15 and 16). These can be explained by the effects and action of these muscles during generation of sound by stretching the syringeal cartilages and narrowing the passages. Increases in the diameter of these muscle are recorded in zebra finch song and Scaup Aythya marila, which is responsible for movements of trachea (Daley and Goller, 2004; Pierko, 2008). Furthermore, the function of these muscles is thought to adjust the length of the trachea during (inspiration and expiration) and control rigidity of the cartilages of the vocal organ (syrinx) during phonation (Burke et al., 2007).
The Triangular voice was triangular shaped space located at the bifurcation of trachea between left and right primary bronchi. The inter-bronchial ligament deemed its base, while the caudal aspect of pessulus composed its apex. Meanwhile, the medial vibrating membranes were formed the lateral borders of the triangle voice (Figure 2). The male had significantly (P˂0.05) greater area than females (Figure 20). Similar Triangular voice shape was found in duck (Frank et al., 2007; Pierko, 2008) and in goose (Onuk et al., 2010).
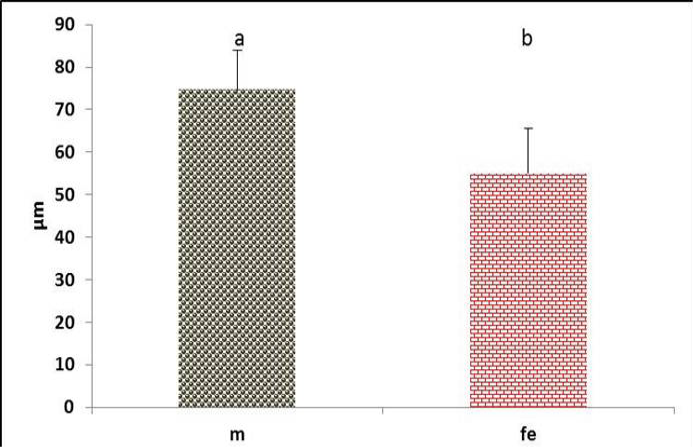
Figure 13: The lateral vibrating membrane thickness in the syrinx of both male and female black francolin. Values represent means±SD. Different letters means significantly (P<0.05) different. Where, male (m); female (fe)
.
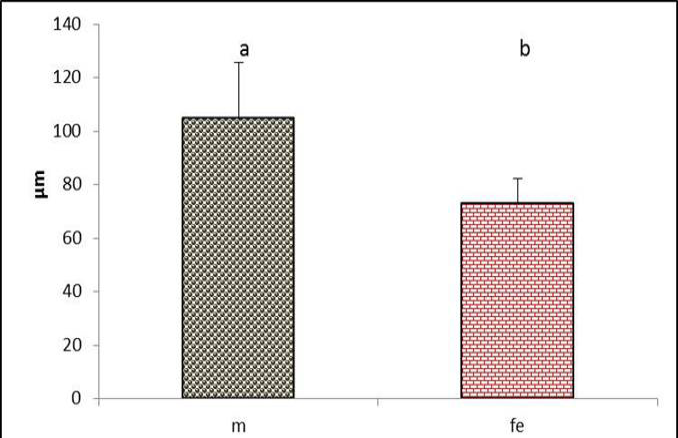
Figure 14: The medial vibrating membrane thickness in the syrinx of both male and female black francolin. Values represent means±SD. Different letters means significantly (P<0.05) different. Where, male (m); female (fe).
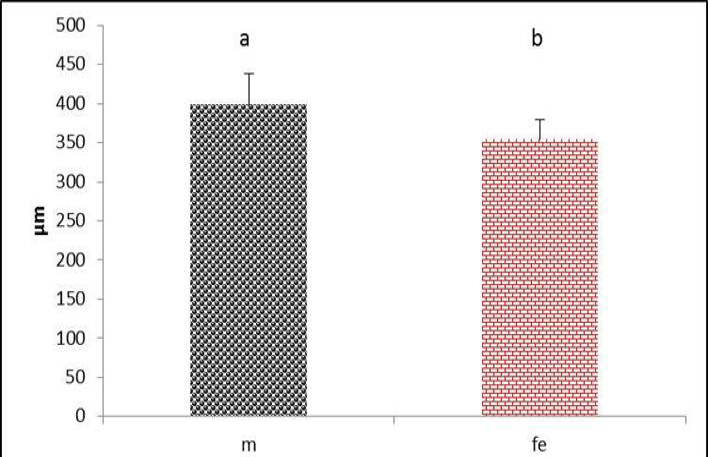
Figure 15 Thickness of sternotrachealis muscle in both male and female black francolin. Values represent means±SD. Different letters means significantly (P<0.05) different. Where, male (m); female (fe).
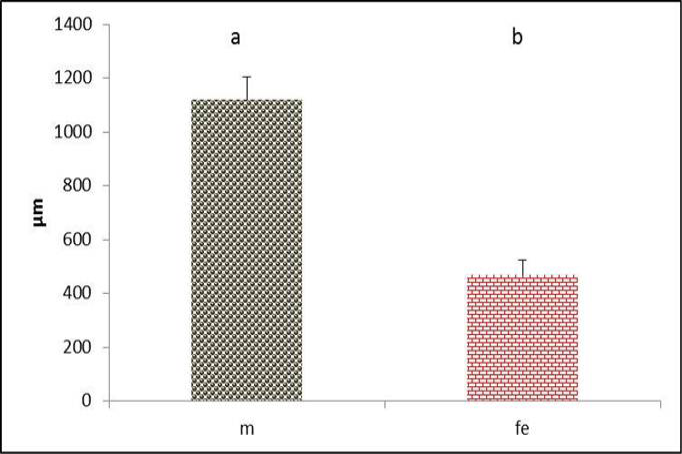
Figure 16: Thickness of lateral trachealis muscle in both male and female black francolin. Values represent means±SD. Different letters means significantly (P<0.05) different. Where, male (m); female (fe). )

Figure 17: Micrograph: The Pessulus (Longitudinal section) of the syrinx in female black francolin. Notice, elongate triangle pessulus showing ossification in most of its parts. Masson´s trichrome stain.

Figure 18: Micrograph: The Pessulus (Longitudinal section) of the syrinx in male black francolin. Notice, short wide triangle pessulus showing ossification in most of its parts. Masson´s trichrome stain.
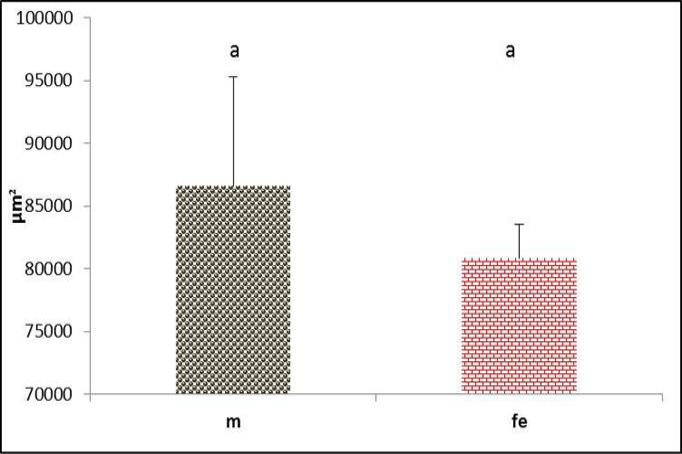
Figure 19: Pessulus area of syrinx in both male and female black francolin. Values represent means±SD. Different letters means significantly (P<0.05) different. Where, male (m); female (fe).
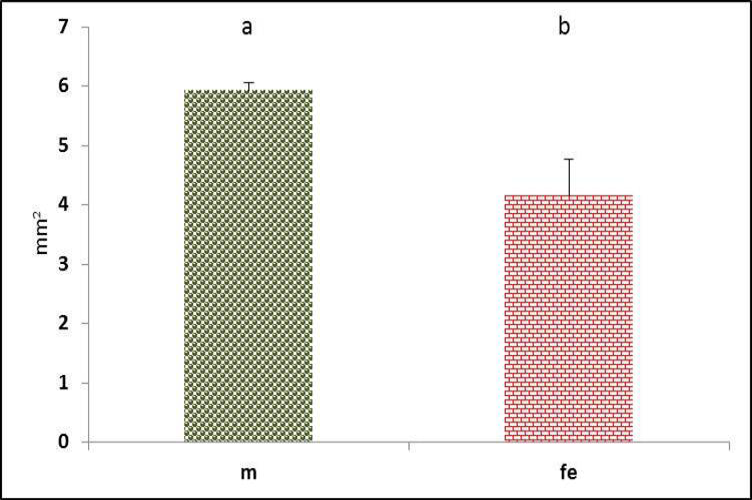
Figure 20: Triangular voice area of syrinx in both male and female black francolin. Values represent means±SD. Different letters means significantly (P<0.05) different. Where, male (m); female (fe).
Through the longitudinal sections of syrinx of black francolin, the pessulus illustrated mostly as ossified cartilage. It had long triangular shaped in female (Figure 17), while it was short triangular in male (Figure 18). Despite that males had greater pessulus area than female but these differences were not significant (P>0.05) (Figure 19). This structure may help to amplify the sound that generated from the syrinx. However, the differences in pessulus shape between genders were mentioned in birds (Aughey and Frye, 2001). It is dagger shaped in domestic fowls (Tasbas et al., 1994), half prism shaped in Japanese quail (Cevik- Demirkan et al., 2007).
In conclusion to the data mentioned above, there was sexual dimorphism which represented generally in male by narrow tympanum and right passage of syrinx in male for more laud sound generation. In addition, greater in syringeal muscles fibers and area of the pessulus and triangular voice that may be magnified the sound wave. The mere presence of sexual dimorphism in these structures may be explained how vocal differences between males and females had arisen.
Conflict of interest
There is no conflict of interest.
Authors’ contribution
Muna-Al aameli: sample collection, lab work, writing; Khalid k kadhim: planning of research, writing, statistical analysis.
Refrences



