Advances in Animal and Veterinary Sciences
Mini Review
Advances in Animal and Veterinary Sciences 1 (4): 102 – 106Zona Free Hamster Oocyte Penetration Test: a Reliable In Vitro Bioassay to Assess the Bull Fertility
Subodh Kumar*, Subuhi Khan, Ayyar Sivakumar, Hirenallur Maheshwarrappa Yathish, Balasubramaniam Sivamani, Deepak Sharma, Arjava Sharma
-
Division of Animal Genetics, IVRI, Izatnagar, Bareilly, Uttar Pradesh, 243 122, India
*Corresponding author:subkum@gmail.com
ARTICLE CITATION:
Kumar S, Khan S, Sivakumar A, Yathish, HM, Sivamani B, Sharma D and Sharma A (2013). Zona free hamster oocyte penetration test: A reliable In Vitro bioassay to assess the bull fertility. Adv. Anim. Vet. Sci. 1 (4): 102 – 106.
Received: 2013–06–30, Revised: 2013–07–02, Accepted: 2013–07–02
The electronic version of this article is the complete one and can be found online at
(
http://www.nexusacademicpublishers.com/table_contents_detail/4/73/html
)
which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited
ABSTRACT
It has been demonstrated that when the zona pellucida of hamster ova was freed, it permitted the entry of spermatozoa from hamster (homologous species) as well as spermatozoa from a variety of species (heterologous species). Suggestions have therefore been made that zona free eggs of golden syrian hamsters (Mesocricetus auratus) may provide a viable alternative to test the penetrability of spermatozoa since they allow entry of capacitated spermatozoa from a wide variety of foreign (heterologous) species. The parameters viz. sperm attached per ovum, ova attached, fertilization percent, fertilization index and polyspermic ova are evaluated in this test. Many workers have used this simple test in laboratory to evaluate the fertility potential of bull under In Vitro conditions. This test have also been used in human to test the penetrability of spermatozoa of the suspected sub fertile man. We developed a simple regression equation at our lab for crossbred cattle and buffalo bulls and found a high correlation between predicted and actual fertility status. This bioassay has proved to have a high association with the fertility status of the livestock species and hence could serve as a useful tool in identifying potentially sub fertile males before they enter into the sire sampling programme. This would tend to make the dairy industry more profitable by cutting down the cost on investment in raising the sub fertile bulls.
INTRODUCTION
The fertilization potential of a dairy bull is an important aspect which contributes significantly to the success of dairy industry. High yielding and meritorious dairy sires might have a lower fertility due to various reasons which would render them less fit in their contribution to genetic improvement. Although fertility is a composite trait and depends upon many biochemical and physiological activities, yet some physical parameters viz. semen volume, progressive motility, sperm concentration, live sperm count, freezability etc. have been applied to predict the fertility status of bull and its conception rate. Beside these, some workers have also used the pedigree record, physical characteristic of bull like growth rate, body conformation and libido as an indirect yardstick to assess the fertility. However, it has been observed that all these parameters do not predict true fertility of the bull (Hafez, 2000) but provide only a rough estimate of the same. Similarly another field based approach used to measure the fertilization potential is the 60–90 days non return rate (NRR). On the other hand some workers have suggested that conception rate (CR) is a better measure than NRR because females, despite being non pregnant, may not return to service due to various physiological abnormalities. Due to high cost of feeds, management and lower socio–economic status of Indian farmers, the bulls are kept to be at the minimum. Even at the organized farms, fewer young males are used for progeny testing and subsequent breeding. This situation calls for a test which would rank the young bulls of progeny testing programme, on the basis of early prediction of their fertilization potential. The laboratory techniques lack accuracy and the field based measures (eg. NRR or CR) take a longer time. Hence there is a strong and genuine need of a technique which would quickly predict the fertilization potential with maximum accuracy.

Many attempts have been made to test the fertilization potential of a sire under the In Vitro system. Yanagimachi (1977) has revealed that hamster oocytes, when zona freed by the action of certain proteolytic enzymes, allow penetration of capacitated spermatozoa from various mammalian species. Many such species that allow penetration of capacitated spermatozoa from various mammalian species have been listed in Table 1. This bioassay may provide a unique tool for evaluating the fertilization potential of spermatozoa under In Vitro conditions. The ideal technique for evaluating fertilizing ability of spermatozoa would be penetration with eggs of homologous species but due to non availability of large number of such eggs, this has less practical importance. The eggs do not allow penetration of sperm from a heterologous species because of sperm eggs interaction in mammals is highly species specific. The zona pellucida of egg forms the major barrier to prevent cross fertilization which can occur with ease if the eggs are zona freed (Yanagimachin, 1977). Hanada & Chang (1972) have demonstrated that when zona of rat ova was freed, it permitted the entry of rat spermatozoa (Homologous species) as well as mouse spermatozoa (Heterologous species). Yanagimachi (1981) has further suggested that zona free eggs of Golden Syrian Hamsters (Mesocricetus auratus) provide a viable alternative since they allow entry of capacitated spermatozoa from a wide variety of foreign (heterologous) species hence offer a reliable penetration test to test the penetrability of spermatozoa.
Zona free hamster eggs are generally used for human beings and have been found to have high correlation with the conception rate of males (Barros et al., 1978) though some workers have found opposite results as well (Zainul Rashid et al., 1998). The eggs also allow the penetration of capacitated spermatozoa of cattle (Bousquet and Brackett, 1982), horse (Brackett et al., 1982), pig (Imai et al., 1977), goat (Shoran & Hanada, 1985), horse (Samper et al., 1989), cat (Howard et al., 1988), poultry (Perkanyi et al., 1992) and off late, buffalo in our laboratory (Kumar and Sharma, 2005). Some workers considered this test to be a complete test to measure the fertilization potential however, Yanagimachi (1984) believes that no single test can measure all the aspects of sperm function. The zona free hamster test measures the ability of spermatozoa to undergo capacitation and thus acrosome reaction, fuse with egg plasma membrane and form sperm pronuclei. It is unable to measure other aspects such as penetration into cervical mucus, surviving in and reaching the site of fertilization and penetrating the cumulus mass and zona pellucida, all of which are essential prerequisites for In Vitro fertilization.
Colonies of Golden Syrian Hamsters (Figure 1) are obtained through systematic breeding. High energy feed and water are supplied ad libitum so as to prevent cannibalism among animals. Hamsters require a high plane of management which includes cleaning of their room regularly with phenyl solution, changing their caging and bedding (rice husk) every 4–5 days, providing them fresh water twice a day and 11–12 hrs of light for breeding purpose. One male is kept with four females and as the females conceive (visualized by fattening), they are put in separate cage. The gestation period is 15–17 days. The off spring are allowed to be with mother up to 21 days. After 8 weeks they are sexed by visualizing external genitalia and kept separately as male and females. About 10–14 week old (100–150 gm body weight) females hamsters are super ovulated to obtain a large number of eggs.

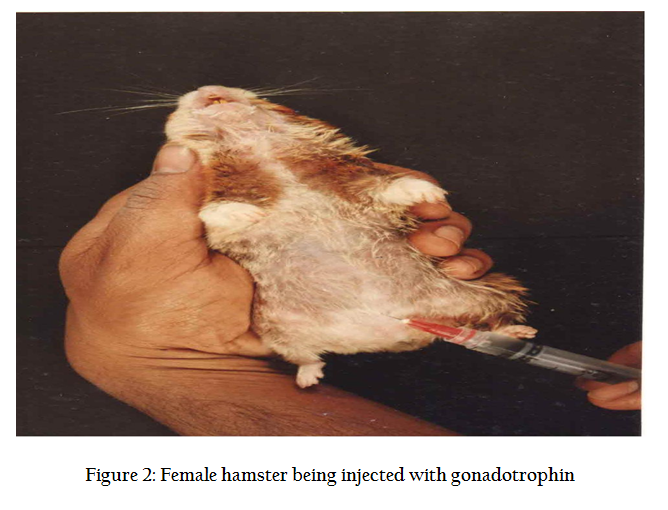
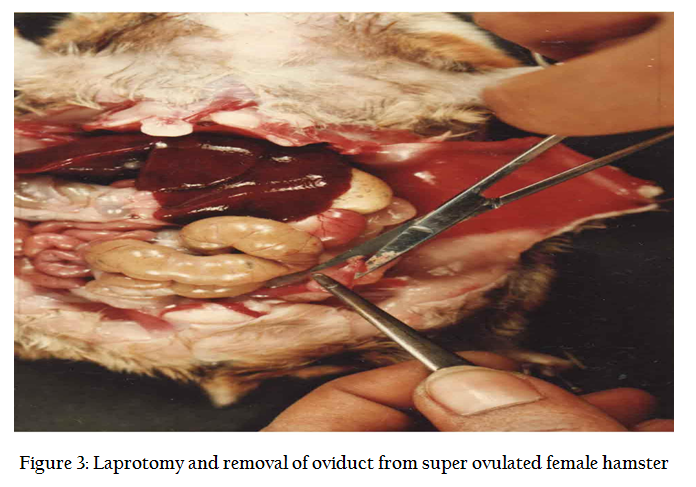
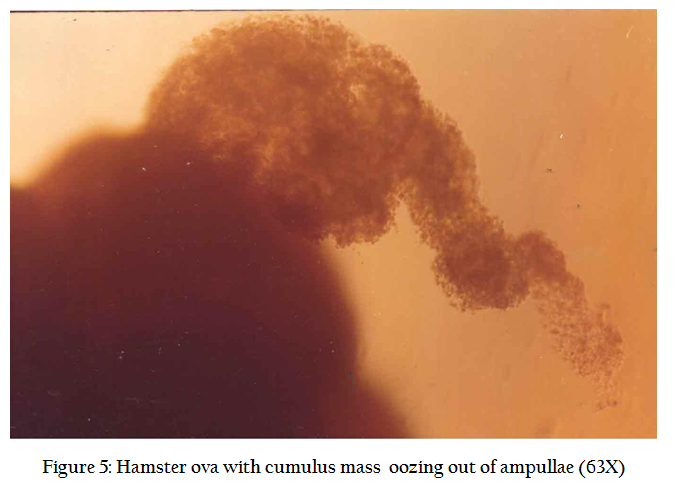
In normal cycle, a female hamster ovulates between 8–16 eggs approx 14 hrs after LH surge. To increase the number of recoverable eggs, super ovulation is induced by intra peritoneal injection (Figure 2) of 50 IU of PMSG (Folligon ® Intervet, Holland) and 98 hrs, another i/p injection of 50 IU of hCG (Chorulon ®, Intervet, Holland). The animals are sacrificed 17–18 hrs after hCG injection by cervical dislocation. Preparation of zona free eggs are performed in the sterile laminar flow hood where all precaution are taken to avoid contamination. Temperature of the room is maintained at 33OC and only red right is used in the room. The animal is pinned down and its musculature is cut open and oviducts are located (Figure 3). Both the oviducts are taken along with little portion of ovary so as to prevent any loss of egg. The cumulus mass is released by pricking the bag like (swollen) region of the ampuellae by a fine forceps (Figure4.) under a dissecting microscope in a clean watch glass containing 1 ml modified sperm TALP medium. Cumulus mass is digested by treating for 1 minutes in 0.1% hyaluronidase enzyme solution. The released eggs (Figure 5) are carefully lifted using self drawn micropipettes attached to a rubber tubing and mouth piece and transferred to a 35 mm disposable petridish having 2 ml sperm TALP medium. The eggs are washed twice so as to ensure removal of all cumulus cells (Figure 6). These are again transferred to 0.1% trypsin solution for 1 minute (Figure 7). From here, eggs are quickly transferred to a 35 mm disposable petri dish having fertilization medium after their zona got freed. Zona free eggs (Figure 8) are once again washed to make them ready for insemination and co–culture. Once the zona free and sperm suspension is ready, a 100 ml drop of sperm suspension (having concentration 3–6 million /ml). 8–10 zona free hamster ova are carefully lifted and gently transferred into sperm suspension drop. The drop is then covered with light mineral oil. The petridishes are kept in a CO2 incubator ser at 35O C with a 5% CO2 level and 99% humidity for 3 hrs. Three hours after the gamete interaction (co–culture), the petri dishes are taken out of incubator. Ova are taken out of sperm suspension drop and washed 4 times in medium. In order to evaluate the penetrability of spermatozoa, ova with small amount of accompanying medium were transferred to a centre of 4 wax spots on a microscopic slide. The ova are separated gently, so that they do not stick to each other. A cover slip was applied over the wax spots and compressed gently in order to flatten them without rupturing.


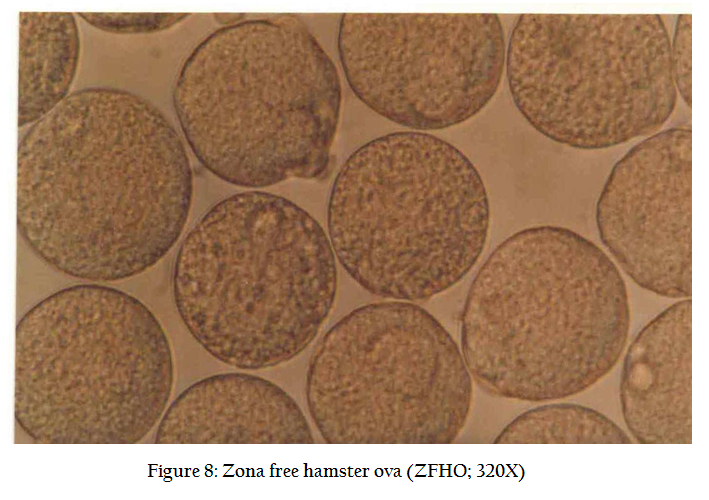
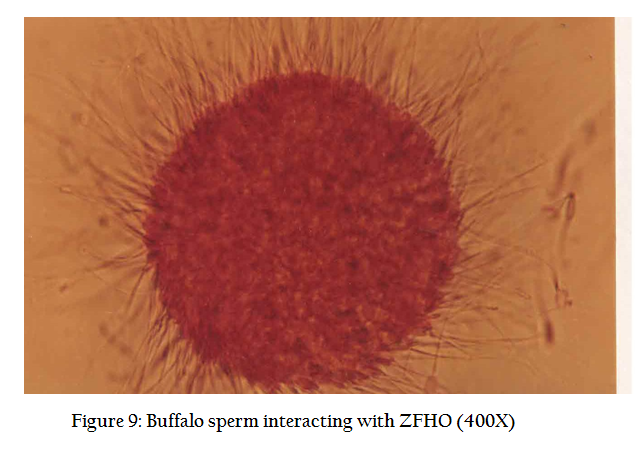
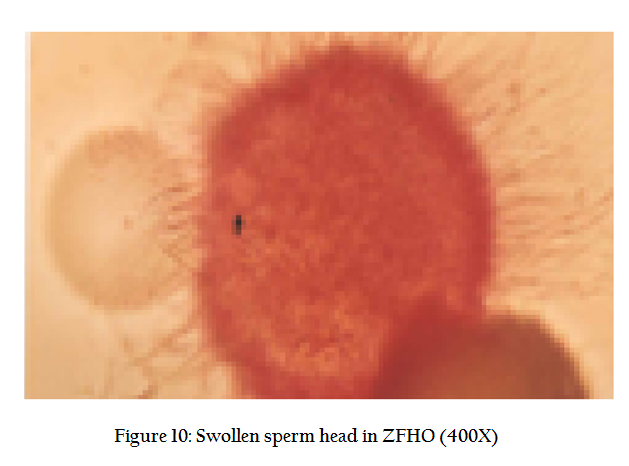
Occasionally before mounting, the ova are stained in 1% eosin stain for a better visualization. We find many spermatozoa stuck to the periphery of ova, vigorously beating their tails and resembling a sunflower (Figure9). A deeper look may show a swollen sperm head that has entered into the ova (Figure 10). The tail of the sperm sheds down and only the sperm head merges into the ovum.
The following parameters are evaluated in zona free hamster oocyte penetration test:
Sperm attached per ovum (SA/O): It is the average number of sperm attached per ovum.
Ova attached (OA): It is the percent ova attached with sperm.
Fertilization percent (FP): It is the percent ova penetrated with sperm (Rogers et al., 1982)
Fertilization Index (FI): It is the average number of sperm heads within per ovum (Rogers et al., 1982).
Polyspermic Ova (PO): It is the percent ova showing poly spermy.
SA/O = (Total number of sperm attached in all the ova)/ (Total number of ova inseminated)
OA (%) = (Total number of ova with attached sperm)/(Total number of ova inseminated) x 100
FP (%) = (Total number of ova penetrated with sperm)/(Total number of ova inseminated) x 100
FI = (Total number of swollen sperm heads within ova)/(Total number of ova inseminated)
PO (%) = (Total number of ova showing poly spermy)/ (Total number of ova inseminated) x 100
Positive trends between sperm vitteli and conception data using frozen semen sample both In Vitro and in vivo were reported by Bousquet and Brackett (1981). High correlation with the percentage of hamster vitelli interacting with frozen thawed semen samples of two Holstein Bull with their 60 days NRR was reported by Bousquet and Braquet (1982). Similar reports were furnished by Berger & Parker (1989) in boar. Capability of zona free hamster eggs to allow penetration of spermatozoa from large number of foreign species does not mean that the former completely lacks the species specificity. The plasma membrane of the hamster egg has the greatest affinity for spermatozoa of its own species. Spermatozoa with intact acrosome are unable to penetrate zona free eggs even though they come in contact with them. Zona free hamster eggs permit penetration only if spermatozoa have undergone both capacitation and acrosome reaction (Yanagimachi, 1981). Further it has been reported that plasma membrane of hamster eggs lack a mechanism to block polyspermy which makes it different from other mammalian species. Employing zona free hamster test, Bousquet et al. (1983) reported the number of sperm interacting per ovum ranged from 1.6 to 3.8, number of sperm attached per ovum ranged from 1.4–2.9, number of sperm penetrated per ovum ranged from 1.5–1.8, percent ova penetrated ranged from 62–85% and percent ova interacting with sperm ranged from 76–92%. The NRR of these bulls ranged from 71.6–75.6%. It was observed that frozen thawed bull spermatozoa treated with 0, 10, 50 & 100 µg/ml heparin, the fertilization percent (% ova penetrated) was 5.4, 50.5, 68.8 and 3.5% and corresponding figure for polyspermy were 0.0, 0.0, 3.0 and 8.0 % respectively (Lu and Gorden, 1988). Thus polyspermy increased with increase in FP. The same phenomenon was reported by Ramesha (1991) who found mean fertilization percent (range) to be 79.49. (59.09–82.61) percent for calcium ionophore A23 187 treated spermatozoa of Karan Fries Bulls.
Recently, Oh et al. (2010) used zona free hamster oocyte and standardized the procedure for the sperm penetration assays, resulting in greatly increased sensitivities for small and large litter size in pigs. The protocol increases the ability to discriminate between good and poor fertility groups and it was highly effective. Park et al. (2012) used zone free hamster test for evaluate and optimize a protocol to determinate bulls according to their sperm fertility ability by sperm penetration assay and found protocol accuracy was 95.7% in both the lower and high NRR with sensitivity of 95.5% and a specificity of 95.8%. Zona free hamster oocytes test was also used to investigate the importance of sperm glutathione treatment in sperm premature chromosome condensation (PCC) by Meybodi et al. (2012). Beside this, several studies on sperm fertility were carried out for human by zona free hamster oocytes penetration test. This test was also used for the assessment of the signaling pathway regulation of zn–α2 glycoprotein in human (Liu et al., 2012), revealing that zn–α2 glycoprotein affect sperm acrosome reaction through both, the cAMP/PKA and PKC pathways and play a critical role in sperm fertility. Other research work on ability of abnormally shaped human spermatozoa to adhere to and penetrate zona free hamster eggs suggest that assessment of morphology may be an unreliable measure for the individual of sperm fertilization ability and emphasized that sperm function testing is an important part of the evaluation of teratospermia (Bronson et al., 2007). Zona free hamster oocyte penetration assay was used to evaluate the possible association between activation of the apoptosis cascade in human sperm and its oocyte penetration capacity results showed that apoptosis–related signaling appears to have a negative association with sperm–oocyte penetration. The exclusion of sperm presenting with those apoptosis–related features during assisted reproduction may improve success rates of procedures such as intrauterine insemination and In Vitro fertilization (Grunewald et al., 2008). The effect of adjudin and different reagents on sperm functions, was evaluated by Li et al. (2013), wherein sperm fertilizing ability was evaluated by sperm penetration of zona–free hamster egg assay. Study demonstrated that adjudin inhibition of capacitation is reversible and its toxicity is low, opening the door for the examination of adjudin as a mediator of male fertility control. Adjudin may be a safe, efficient and reversible male antifertility agent and applicable to initial clinical trials of adjudin as a male anti fertility agent in humans.
We have suggested a linear predictive equation in cattle and buffalo, to predict the fertility potential of bulls in terms of conception rate by employing the ZFHOPT. We performed zona free hamster oocyte penetration test on crossbred cattle and buffalo. We deliberately chose the crossbred cattle because sperm motility and fertility have known to be a problem in crossbred bulls and their rejection rate have reported to be as high as 50% (Geetha et al., 2011). We found that, the average sperm attached per ovum in uncapacitated (controls) and capacitated (treated with capacitating agent) spermatozoa samples were 14.22 ± 0.52 and 20.47 ± 0.61; 126.48 ± 2.09 and 145.38 ± 1.86, for crossbred cattle and buffalo spermatozoa respectively. The ova attached for the same averaged 94.79 ± 1.02 and 97.72 ± 0.67 % for crossbred cattle spermatozoa whereas for buffalo spermatozoa, the values for both, controls and treated samples averaged 100.00% (net increase being 0.00%). The mean fertilization percent for controls and treated cattle spermatozoa were 54.21 ± 1.98 and 74.16 ± 1.42 % and the corresponding values for buffalo spermatozoa were 74.21 ± 1.59 and 89.11 ± 1.18 % respectively. The corresponding values for fertilization index averaged 0.55 ± 0.02 and 0.78 ± 0.02 and 0.79 ± 0.02 and 1.10 ± 0.03 for crossbred cattle and buffalo spermatozoa respectively. The same values for polyspermic ova averaged 0.00 ± 0.00 and 4.48 ± 0.99 and 5.22 ± 1.22 and 21.69 ± 1.88 % for crossbred cattle and buffalo spermatozoa respectively. The increase in sperm attached per ovum, fertilization percent, fertilization index, due to PAF treatment was significant (P< 0.01) in both the species whereas it was significant (P<0.05 ) for ova attached in cattle and polyspermic ova ( P <0.05 in cattle and <0.01 in buffalo). The fertilization parameters, viz. the sperm attached per ovum and fertilization index (for treated samples) in both the species had the highest R2 values. These values were jointly regressed against the observed conception rates and the values of intercept constant (A), regression coefficients (B1 and B2) were calculated. A prediction equation (Y= A+B1X1 + B2X2) employing these two variables was fitted and the conception rates (CR) were predicted. The predicted CR showed a significant (P<0.01) sample coefficient (r) and rank correlation coefficient (R) with the observed CR in both the species viz. crossbred cattle and buffalo.
CONCLUSION
As because the results in our laboratory indicated that the ZFHOPT exhibited a high correlation between predicted and the actual fertility, it could hence be concluded that ZFHOPT could serve as a useful tool in identifying potentially sub fertile males before they enter into the sire sampling programme and make the dairy industry more profitable by cutting down the cost on investment in raising the sub fertile males.
REFERENCES
Barros C, Gonzalez J, Herrera Eand, Bustos–Obregon E (1978). Fertilization capacity of Human spermatozoa evaluated by actual penetration of foreign eggs. Contracept. 17: 87–92.
http://dx.doi.org/10.1016/0010-7824(78)90064-1
Berger T and Parker K (1989). Modification of zona free hamster ova bioassay of boar sperm motility and correlation with in vivo fertility. Gam. Res. 22 (4): 385–397.
http://dx.doi.org/10.1002/mrd.1120220405
PMid:2722120
Bronson RA, Bronson SK and Oula LD (2007). Ability of abnormally shaped human spermatozoa to adhere to and penetrate zona free hamster eggs: Correlation with sperm morphology and post–incubation motility. J. Androl. 28(5): 698–705
http://dx.doi.org/10.2164/jandrol.106.001503
PMid:17460097
Bousquet D and Brackett BG (1981). Penetration of zona free hamster ova by bull sperm after frozen storage. Therio. 15 (1):117–123.
http://dx.doi.org/10.1016/S0093-691X(81)80034-9
Bousquet D and Brackett BG (1982). Penetration of zona free hamster ova as a test to assess fertilizing ability of bull sperm after frozen storage. Therio. 17 (2):199–213.
http://dx.doi.org/10.1016/0093-691X(82)90081-4
Bousquet D, Bracket BG, Dressel MA and Allen CH (1983). Efforts to correlate laboratory with field observations on bull sperm fertility. Therio. 20 (5): 601–613.
http://dx.doi.org/10.1016/0093-691X(83)90084-5
Brackett BG, Bousquet D, Boice ML, Donawick WJ, Ewans JF and Dressel MA (1982). Normal development following in vitro fertilization in cow. Biol.Reprod. 27 (1):147–158.
http://dx.doi.org/10.1095/biolreprod27.1.147
PMid:6896830
Geetha T, Kumar S, Dubey PP, Sivamani B, Ghosh SK, Mitra A, Tomar AKS and Sharma A (2011). Sequence variability in CatSper1 gene in Vrindavani crossbred cattle. Ind. J. Anim. Sci. 81 (9): 981–983.
Grunewald S, Said TM, Paasch U, Glander, HJ and Agarwal A (2008). Relationship between sperm apoptosis signaling and oocyte penetration capacity. Int J Androl. 31 (3): 325–330.
http://dx.doi.org/10.1111/j.1365-2605.2007.00768.x
PMid:17573851
Hafez ESE (2000). Semen evaluation. In Reproduction in farm animals (7th edition, ed. ESE Hafez), Lea and Febiger, Philadelphia, U.S.A.
Hanada A and Chang MC (1972). Penetration of zona free eggs by spermatozoa of different species. Biol. Reprod. 6: 300–309.
PMid:4552741
Howard JG, Barone MA, Clingerman KJ, Bush M and Wildt DE (1988). Influence of teratospermia, culture media and a sperm swim up tecjnique on penetration of zona free hamster oocytes by domestic cat sperm. In 11th Int. Congr. Anim. Reprod.artif. Insemin., paper no. 332, 3 pp, University College, Dublin, Ireland, June 26–30, 1988.
Imai H, Niwa K and Iritani A (1977). Penetration in vitro of zona free hamster eggs by ejaculated boar spermatozoa. J. Reprod. Fert. 51: 495–497.
http://dx.doi.org/10.1530/jrf.0.0510495
PMid:592305
Jackubicka I, Barta M and Babusik P (1989). Fertilizing capacity of the ejaculated sperm in nutrias (Myocastor coypus) following extirpation of seminal vescicles, determined by penetration test and natural mating. Vet. Med. 34 (7): 441–446.
Kumar S and Sharma A (2001). Prediction of fertilization potential of frozen spermatozoa of cattle and buffalo. Ind J Anim Sci 71 (4): 331–333.
Kumar S and Sharma A (2005). Platelet activating factor improves the in vitro penetration of zona free hamster eggs by buffalo (Bubalus bubalis) spermatozoa. Therio 63: 1564–1572.
http://dx.doi.org/10.1016/j.theriogenology.2004.05.028
PMid:15763101
Li K, Ni Y, He Y, Chen WY, Lu JX, Cheng CY, Ge RS and Shi QX (2013). Inhibition of sperm capacitation and fertilizing capacity by adjudin is mediated by chloride and its channels in humans. Hum Reprod. 28 (1): 47–59.
http://dx.doi.org/10.1093/humrep/des384
PMid:23117128 PMCid:PMC3522416
Liu Y, Qu F, Cao X, Chen G, Guo Q, Ying X, Guo W, Lu L and Ding Z (2012). Con A–binding protein Zn–α2–glycoprotein on human sperm membrane is related to acrosome reaction and sperm fertility. Int J Androl. 35 (2): 145–157.
http://dx.doi.org/10.1111/j.1365-2605.2011.01195.x
PMid:21790656
Lu KH and Gordon I (1988). Effect of heparin on the capacitation of frozen thawed bovine spermatozoa used in the in vitro fertilization (IVF) of mouse oocytes matured in vitro. In 11th Int. Cong. Anim. Reprod. Artif. Insemin., paper number : 339, University College, Dublin, Ireland, June 26–30,1988.
Meybodi AM, Mozdarani H, Moradi SZ and Akhoond MR (2012). Importance of sperm glutathione treatment in ART. J Assist Reprod Genet. 29 (7): 625–630
http://dx.doi.org/10.1007/s10815-012-9755-5
PMid:22492222 PMCid:PMC3401252
Oh SA, You YA, Park YJ and Pang MG (2010). The sperm penetration assay predicts the litter size in pigs. Int. J. Androl. 33 (4): 604–612.
PMid:19538520
Park YJ, Mohamed–el SA, Oh SA, Yoon SJ, Kwon WS, Kim HR, Lee MS, Lee K and Pang MG (2012). Sperm penetration assay as an indicator of bull fertility. J. Reprod. Dev. 58 (4): 461–466.
http://dx.doi.org/10.1262/jrd.11-067H
PMid:22522227
Pavlok A, Petelikova J and Flechon JE (1983). Interaction of zona free hanster eggs with fresh and frozen ram spermatozoa in vitro. In The Sperm Cell (J.Andre–Martinus, ed.), pp:51–54, Nijhoff, The Hague.
http://dx.doi.org/10.1007/978-94-009-7675-7_10
Perkanyi V, Babusik P, Bednarczyk M and Baumgartner J(1992). A technique to examine cock sperm chromosomes using zona free hamster eggs. Folia. Biol. Praha., 38 (1): 25–30.
Ramesha KP (1991). Studies on sperm chromosome of bovines. Ph.D.Thesis (submitted to N.D.R.I., Karnal).
Rogers BJ, McCarville C, Soderdahl D and Hale R (1982). Re –evaluation of the zona free egg test with regard to its use in human fertility assessment. Fertil. Steril. 37: 296–301.
Samper JC, Behnke EJ, Byers AP, Hunter AG and Crabo BG (1989). In vitro capacitation of stallion spermatozoa in calcium free Tyrodes medium and preparation of zona free hamster eggs. Therio. 31 (4): 875–884.
http://dx.doi.org/10.1016/0093-691X(89)90032-0
Shoran B and Hanada A (1985). Penetration of zona free hamster eggs in vitro by ejaculated goat spermatozoa after treatment with ionophore A 23187. Jap. J. Anim. Reprod. 31: 115–121.
http://dx.doi.org/10.1262/jrd1977.31.115
Takahashi Y, Nihayah M, Hishinuma M, Jainudeen MR, Mazni, AO, Mory Y and Kanagawa H (1989). Preliminary study of buffalo sperm penetration into zona free hamster eggs after treatment with calcium ionophore A 23187. Jap. J. Vet. Res. 37: 161–166.
PMid:2515337
Yanagimachi R (1977). Specificity of sperm egg interaction. In Immunobiology of Gametes (MH Johnson, ed.),pp: 225–295, Cambridge University Press, London.
Yanagimachi R (1981). Mechanism of fertilization in mammals.In Fertilization and embryonic development in vitro (L Mastroianni and JD Biggers, eds.), pp:81–82, Plenum Press, New York.
http://dx.doi.org/10.1007/978-1-4684-4016-4_6
Yanagimachi R (1984). Zona free hamster eggs: their use in assessing fertilizing capacity and examining chromosomes of human spermatozoa. Gam. Res.10: 187–232.
http://dx.doi.org/10.1002/mrd.1120100210
Zainul Rashid MR, Fishel SB, Thornton S, Hall JA, Ndukwe G, Aloum M and Fleming SD (1998). The predictive value of the zona–free hamster egg penetration test in relation to in–vitro fertilization at various insemination concentrations. Hum. Reprod. 13 (3): 624–629.
http://dx.doi.org/10.1093/humrep/13.3.624
PMid:9572423



