Advances in Animal and Veterinary Sciences
Short Communication
Advances in Animal and Veterinary Sciences 1 (2): 69–74Characterization of Virulence Factors among Diverse Salmonella Serotypes and Sources
Shweta Singh1, Himanshu Singh2, Suresh Chandra Tiwari3, Prejit Nambiar4, Rajesh Kumar Agarwal1
- National Salmonella Centre (Vet), Division of Bacteriology and Mycology, Indian Veterinary Research Institute, Izatnagar, Bareilly, 243122, U.P. India
- Department of Biotechnology, Lovely Professional University, Jalandhar, India
- Department of Botany and Microbiology, Hemwati Nandan Bahuguna Garhwal Central University, Srinagar, Uttrakhand, India
- Department of Veterinary Public Health, College of Veterinary and Animal Sciences, Pookode, Wayanad, India
*Corresponding author: grace_bly@yahoo.com
ARTICLE CITATION:
Singh S, Singh H, Tewari SC, Prejit N, Agarwal RK (2013). Characterization of virulence factors among diverse Salmonella serotypes and sources. Adv. Anim. Vet. Sci. 1(2): 69–74.
Received: 2013-04-01, Revised: 2013-06-02, Accepted: 2013-06-24
The electronic version of this article is the complete one and can be found online at
(
http://www.nexusacademicpublishers.com/table_contents_detail/4/52/html
)
which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited
ABSTRACT
A total of 37 Salmonella isolates of 11 different serotypes and rough type from human and animal clinical cases and meat samples were studied for the presence of 8 virulence determinants including 4 virulence genes and 4 toxic factors. All the Salmonella isolates harboured invA and stn genes, whereas 94.59% of isolates had the presence of fimH and hilA genes indicating colonizing, invasive and enterotoxic potential of the pathogen. Among the four toxic factors studied, highest percentages of the isolates were positive for cytotoxicity (72.97%), followed by DNAse (70.27%), enterotoxic (51.35%) and hemolysin (13.51%) activities. All the isolates were found to be highly pathogenic as indicated by 100% mortality in mouse model. Among the different serotypes studied, Typhimurium and Paratyphi B var Java showed the presence of all the virulence determinants in varying degree. The 2 rough strains demonstrated least number of virulence determinants. Tshiongwe and Labadi serotypes were also devoid of enterotoxigenic, DNAse and hemolytic activities. Other serotypes also lacked 1-2 virulence factors. Salmonella isolates exhibited diverse virulence determinants and were highly pathogenic.
Salmonellosis is a major infectious disease of humans and animals globally. Salmonella infections cause heavy economic losses in United States amounting to $ 2-4 billion due to loss of life, work and medical care (Buzby et al., 1996; Frenzen et al., 1999). Different disease syndromes of salmonellosis are gastroenteritis, typhoid, bacteremia, and local infections (Darwin and Miller 1999). Nontyphoidal salmonellosis is manifested by mild to moderate gastroenteritis (CDC 2001). In very few casesa small percentage of cases, septicemia and invasive infections of organs and tissues can occur, which may result in diseases such as pneumonia, osteomyelitis, and meningitis (Cohen et al., 1987).
The ability of Salmonella spp. to cause a disease largely relies on a repertoire of elaborate virulence determinants including toxins fimbriae, flagella and virulence-plasmids (Van Asten and Van Dijk 2005). Salmonella produces both endotoxins and exotoxins (Houston et al. 1981, Ray et al. 1987). The endotoxin is lipid A of the outer membrane lipopolysaccharide (LPS) of Salmonella. The exotoxins are of two type viz., cytotoxins and the enterotoxins. Differences in virulence among Salmonella serovars have been attributed to the variable acquisition and evolvement of virulence genes (Falkow 1996). Several Salmonella specific virulence genes which takes an important role in the pathogenicity have been identified (Baumler et al., 2000). In Typhimurium serovar, at least 80 different virulence genes have been identified. Some genes are known to be involved in adhesion and invasion, like sef (Clouthier et al., 1993), FimH (Duncan et al., 2005), invA (Galan et al., 1992) and other genes associated with toxin production viz., stn (Makino et al., 1999).
The present study was undertaken to determine some of the important virulence factors of Salmonella isolated from human, animals and meats.
A total of 37 Salmonella belonging to 11 serovars (Paratyphi B var java - 14, Typhimurium - 8, Enteritidis - 3, Isangi - 3, Virchow -1, Heidelberg -1, Berta -1, Tshiongwe -1, Saintpaul -1, Bsilla -1, Labadi -1 and 2 rough strains isolated from human and animal clinical cases and meat samples were included in the study. The isolates were confirmed through biochemical, followed by serological tests (Agarwal et al., 2003, Farmer 1995) and maintained on nutrient agar slants.
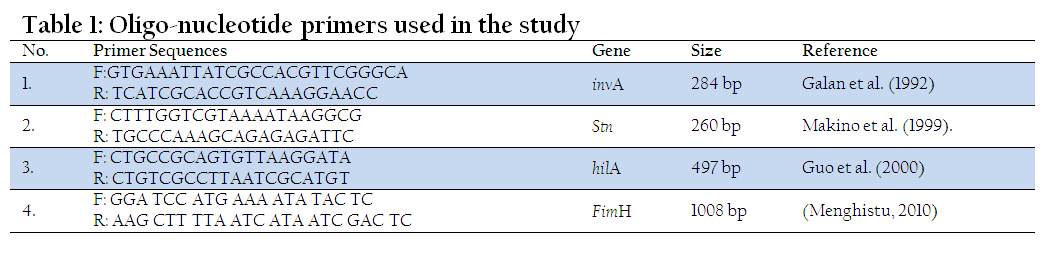
Primers used in the study were custom synthesized from Genuine Chemical Corporation (GCC), New Delhi (Table 1) for the detection of various virulence genes.
All the isolates were grown in brain heart infusion broth at 37 0C for 18 hrs. One ml of BHI grown sample was drawn and subjected to template DNA preparation by boiling and snap chilling method (Singh et al., 2009). Briefly, cultures were centrifuged at 10000 rpm for 2 min at room temperature, collected pellet was dissolved in 200 µl distilled water and put in boiling water bath for 10 min and chilled immediately on crushed ice for 10 min. For PCR 5 µl of supernatant was used as template from this preparation.
All the isolates of Salmonella were subjected to PCR for the detection of invA (Galan et al., 1992), hilA (Guo et al., 2000), fimH (Menghistu, 2010) and stn (Makino et al., 1999) genes. The reaction mix invariably consisted of 5 µl of the bacterial lysate, 5 µl of 10x assay buffer for Taq polymerase containing 1.5 mM MgCl2, 2 µl of 10mM dNTP mix 1 µl each of forward and reverse primer (10 pmol) and 1.25 U of Taq DNA polymerase made upto 50 µl using sterile distilled water. The optimal cycling conditions used for different genes are mentioned in Table 1. At the end of reaction the amplified product was electrophoresed on agarose gel (1.5%), visualized under UV light.
Cell free supernatant of Salmonella isolates was prepared as per Agarwal, 1997. The isolates were grown in brain heart infusion (BHI) broth fortified with 3% (w/v) cassamino acid and 0.6% (w/v) yeast extract, incubated in a shaker (200 rpm) at 370C for 12 h, centrifuged at 10000 g for 30 min at 40C and supernatant was collected. The cell free supernatants (CFS) were filtered through 0.22 µm filters and stored at -200C till further use.
Cytotoxic potential of the isolates were tested on African green monkey kidney cells (vero) lines grown in minimal essential medium (MEM) with Eagle's salts containing 10% fetal bovine serum, 100 U of benzylpenicillin/ml, 100 μg of streptomycin/ml, amphotericin-B 100 μg/ml, 2 mM L-glutamine and 150 μg of G 418/ml.
The Vero cells were grown to confluent monolayer in 96 well tissue culture plate (Greiner, Germany). Two fold dilution of each CFS were prepared and inoculated in triplicate in separate wells, incubated at 370C in 5% CO2 atmosphere and the cytopathic changes (CPE) were recorded at different intervals up to 48 h PI.
The enterotoxigenic activity was determined by vascular permeability reaction (VPR) assay according to the procedure of Sandefur and Peterson (1976).The toxin preparations were injected intra dermal in 0.1 ml quantity in rabbit. After 2 and 18 h of inoculation for rapid permeability factor (RPF) and delayed permeability factor (DPF), respectively. The skin at the site of inoculation was examined for erythema, induration, necrosis and any other changes.
Mouse lethality assay was carried out as per Agarwal et al. (2003). Mice weighing about 16-20 g was injected with 0.2 ml of 20 h broth culture (approx <107 cells) of test organism intra-peritonealy and observed for 5-7 days for the death of the mouse. Each isolate was injected in to 4 mice. The organism was then recovered from liver, spleen and heart blood. Gross pathological lesions were also recorded.
DNAse activity of all the Salmonella isolates was studied on DNAse test agar (Himedia, Mumbai) as per Kavitha et al., (2007). After spot inoculation plates were incubated at 37oC for 48 h to observe clearance around colonies for the detection of extracellular DNAse activity. Plates were exposed to ultraviolet rays (150 to 3900Ao) for half an hour and incubated at 37oC for 24 h. A strain of Staph aureus was included as positive control, which consistently produced purple zone around the colonies.
The haemolysin production was determined by inoculating the sheep blood agar plates by stab method to create an anaerobic pocket. After incubation for 48 h at 370C, the plates were examined for any discoloration or haemolysis and kept at 40C for hot and cold lysis activity, if any (Okazaki et al. 2003).
Thirty seven isolates from human, animal and meat sources were characterized for the presence of 4 virulence determining genes (invA, hilA, fimH and stn) and 4 phenotypic virulence characteristics viz. (cytotoxicity, entertoxigenicity, hemolysin and DNaes activity). Screening of 37 Salmonella isolates for fimH and hilA gene revealed 35 (94.597%) to be positive (Table 2 and 3). Two isolates negative for both these genes were of rough type and were isolated from animal diarrheal cases. All the isolates were positive for invA and stn genes.
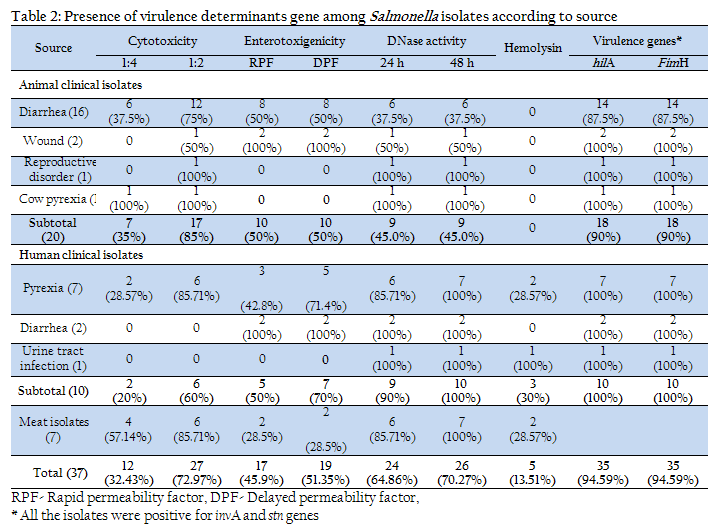
Enterotoxic potential of Salmonella isolates was carried out by vascular permeability assay in rabbit skin. Both rapid permeability factor (RPF) and delayed permeability factor (DPF) were observed. The test revealed that of the 37 Salmonella isolates, only 17 (45.9%) and 19 (51.35%), respectively, showed RPF and DPF activity. The positive reaction was indicated by haemorrhagic zone of varying degree and in some cases enduration and necrosis. Among the different clinical conditions, 100% isolates from human diarrhea and cow wound followed by human pyrexia (71.4%) and animal diarrhea (50%) cases were positive for the enerotoxic activity (Table 2). Furthermore, only 2 (28.5%) of meat isolates were found to be enterotoxigenic. Analysis according to serotype indicated that none of the isolates belonging to Enteritidis, Berta, Saintpaul, Labadi and Virchow serotypes were positive for VPR. Other serotypes produced variable reaction (Table 3).
Cytotoxic potential of all the 37 Salmonella isolates were studied on Vero cell line. Microscopic examination of different cells following exposure to CFS (1:2 and 1:4 dilution) prepared from Salmonella isolates revealed number of changes including rounding and shriveling of cells, loss of cytoplasmic membrane, extension, disorganization of cell sheets, shrinking of cytoplasm etc. Of the 37 isolates tested, 27 (72.97%) isolates were found to be cytotoxic in nature at 1:2 dilutions, whereas at 1:4 dilutions only 12 (32.43%) isolates were cytotoxic (Table 2). Highest (85.71%) cytotoxic potential was demonstrated by isolates of meat, followed by the isolates of animal clinical origin (75%) and human clinical (60%) origins. Among different clinical cases, highest cytotoxic activity was shown by isolates of reproductive disorder and cow pyrexia origin (100% each), followed by human pyrexia (85.71%) and wound (50%) isolates. The 2 isolates from human diarrhea cases and 1 UTI case did not demonstrate any cytotoxic potential.
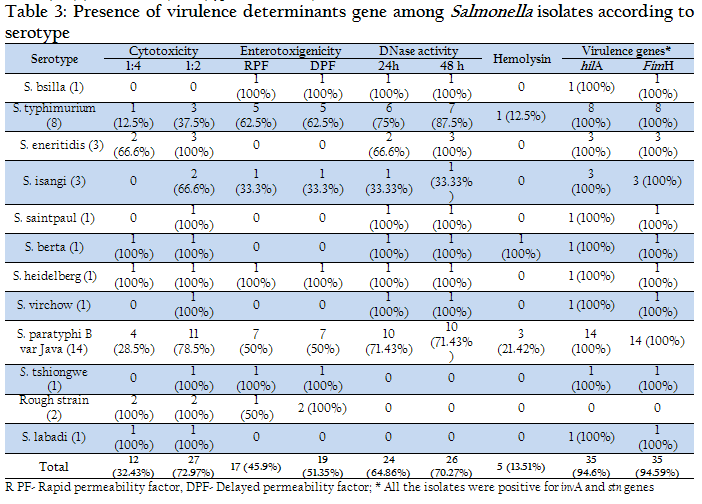
CSF of all the isolates of serotypes Eneritidis, Saintpaul, Berta, Heidelberg, Virchow, Tschicngwe, Labadi, as well as Rough strains were found to be cytotoxic at 1:2 dilution, however, a few of these strains were not toxic at 1:4 dilution. Interestingly, only 11 of 14 isolates of S. Paratyphi B var Java, 2 of 3 isolates of Isangi and 4 of 8 (50%) of S. Typhimurium serotypes were cytotoxic at 1:2 dilution (Table 3).
Screening of all the 37 isolates on sheep blood agar for hemolytic potential revealed that only 5 (13.51%) isolates were positive for β hemolysis. Two of the isolates were from human pyrexia cases, 1 from UTI and 2 were from meat. Moreover, 3 of the positive isolates were Paratyphi B var Java and 1 each were Typhimurium and Berta serotypes.
Examination for DNAse activity of all the Salmonella isolates indicated that after 24 h of incubation 24 (64.86%) of the 37 strains were positive. After 48 h of incubation, 2 more strains i.e. a total of 26 (70.27%) strains turned out to be positive. On further incubation no change in the size of zone or in the number of strains giving positive reaction was seen. Among the different sources of the isolates, only 6 of 16 isolates from animal diarrheal cases and 1 of 2 isolates from wounds produced positive reaction. The DNAse negative strains belonged to S. Paratyphi B var Java (4/14), S. Typhimurium (1/8), S. Isangi (2/3), S. Tschicngwe (1/1), S. Labadi (1/1) serotypes and rough type (2/2).
Mouse pathogenicity assay revealed 100% mortality for all the 37 isolates within 24 h. The organisms were re-isolated from liver, spleen and heart of all the dead mice. On gross examination visceral organs were found to be highly congested, there were hemorrhages in intestine and fluid accumulation in peritoneal cavity.
Salmonella are able to colonize multiple sites including the small intestine, colon, and cecum. Intestinal adhesion in Salmonella infection is mediated by fimbriae present on the bacterial cell surface. Type 1 fimbriae (Fim), thin aggregative, long polar fimbriae (Lpf), and plasmid-encoded fimbriae (Pef) are some among the many types of fimbriae that are involved in Salmonella colonization (Darwin and Miller 1999). FimA, and FimH are the major structural subunit of the 7 genes (fimAICDHF) of Type 1 fimbriae, which interacts with host cell to facilitate attachment (Darwin and Miller 1999; Duncan et al., 2005). FimH gene was selected in this study to assess the ability of Salmonella isolates to colonize the intestine as it could be critical to microbial pathogenesis (Duncan et al., 2005). All the isolates of Salmonella in the study except the two rough strains were found to posses fimH gene (Table 2 and 3) indicating their ability to colonize the intestine.
Invasiveness is the next step in Salmonella pathogenesis determined by several genes located in Salmonella pathogenicity island 1 (SPI1) (Mills et al., 1995). HilA (hyperinvasive locus) is one of the virulence genes located in SPI1. (Bajaj et al., 1995). This study revealed 94.59% isolates to be positive. The 2 negative isolates were of rough type and were isolated from animal diarrheal cases.
Guo et al. (2000) tested 83 Salmonella strains of different serotypes and found all of them to be positive for this gene. However, their study did not include rough variants, which were negative in this study. Pathmanathan et al., (2003) also reported its presence in a total of 33 Salmonella strains of 27 serovars. In an interesting study, Hu et al., (2008) reported that 2 Salmonella isolates of serovar Senftenberg from human clinical cases lacked some genes including hilA critical to Salmonella pathogenicity island 1 (SPI-1). Absence of hilA gene has also been reported in Orion serovar (Trafny et al., 2006). Thus our study supports the fact that some strains of Salmonella may not posses’ hilA gene.
We also examined all the isolates for the presence of invA gene, which is also important for entry into the intestinal mucosa. All the isolates were found to be positive for this gene. The result is in conformity to earlier reports (Galan et al., 1991; Swamy et al. 1996; Abouzeed et al., 2000).
Cytotoxic and enterotoxic activities in Salmonellae have been described (O’Brien and Holmes 1996). Salmonella induced diarrhoea is a complex phenomenon involving several oxins. Enterotoxin, a 29-kDa protein encoded by stn gene is most commonly studied (Chary et al., 1993). This stn gene has been observed in all Salmonella serovars investigated except S. bongori (Prager et al., 1995). The present report also found stn gene in all the 37 isolates studied. Murugkar et al., (2003) studied the distribution of stn gene in 95 Salmonella isolates from five different serovars and four different sources and found that it was present in all the isolates. Rahman (1999) also reported the prevalence of Salmonella enterotoxin (stn) gene in all the 26 strains of Salmonella enterica studied by him. It is evident from our study that stn gene is commonly found among different Salmonella serovars.
It is suggested that enterotoxigenic properties are vital for pathogenesis of Salmonella diarrhea. Vascular permeability reaction in rabbit skin has been shown to demonstrate enterotoxigenic potential of Salmonella by several workers (Kaura et al. 1982; Chary et al., 1993). Culture filtrates of Salmonella have been shown to produce two skin permeability factors (PF). A heat stable rapid acting factor causes bluing of the skin. The second one is a heat labile delayed factor, which results in marked indurations of the rabbit skin within 18 h (Sandefur and Peterson 1976). Chary et al. (1993) demonstrated enterotoxic activity by VPR altered vascular permeability in rabbit skin of a 25 kDa protein expressed from stn gene. In this study, 45.9% and 51.35% isolates showed RPF and DPF activity, respectively. Earlier reports have shown that 55% to 72% (Kuhn et al., 1978; Jiwa 1981) of Salmonella isolates to produce enterotoxin. Slightly lower enterotoxigenic activity recorded in our study may be due to variations in strain, source of isolation or bioassay model used. It is also interesting to note that all the isolates were positive for stn gene but less than half produced enterotoxin. Thus, it appears that mere presence of gene may not necessarily result in enterotoxin production and even stn mutants of S. Dublin and S. Typhimurium in comparision to wild-type strains shows no difference in fluid secretion (Watson et al., 1998). Another interesting finding was that only 50% of animal diarrheal isolates were positive for enterotoxic activity. This indicated that there might be some other mechanism involved in diarrheal pathogenesis (Giannella et al., 1975). Moreover, Wallis et al., (1986) found no correlation between enterotoxigenicity in vitro and the ability of the organisms to produce disease in vivo. Some of the isolates from other clinical conditions such as cow wound (100%), human pyrexia (71.4%) and as well as meat isolates (28.5%) were also found to be enterotoxigenic. None of the isolates belonging to Enteritidis, Berta, Saintpaul, Labadi and Virchow serotypes were positive for VPR. Other serotypes produced variable reactions (Table 2).
In recent years cytotoxin production by enteric pathogens has been studied. These toxins are defined by their ability to kill mammalian cells (van Asten and van Dijk 2005). In this study, overall 72.97% isolates were found to be cytotoxic. The cytotoxicity ranged from 60% to 85.71% in isolates from diverse origin. Among different serotypes studied, serotypes Eneritidis, Saintpaul, Berta, Heidelberg, Virchow, Tshiongwe, Labadi and Rough strains were found to be cytotoxic. Several serotypes including Typhimurium, Enteritidis Choleraesuis have been shown to have cytotoxic potential (O'Brien et al. 1982; Ketyi et al., 1979; Chiu and Ou 1996; Prager et al., 1995; Baloda et al., 1983), but the amount of toxin produced by the different serovars may vary (Ashkenazi et al., 1988).
In search of other virulence factors, all the isolates were also screened for hemolysin and DNAse activities. Hemolysin is one of the virulence factors identified in Salmonella, which is useful in invasion of epithelial cell and cell-surface factor induction It also helps in survival in macrophages and the counteraction of host immune defenses (Uppal et al., 1998). Although slyA gene encoding for hemolysin is present in all Salmonella, the organism is not usually hemolytic on culture media. In low-passage clinical isolates, weak hemolysis may be occasionally observed (Libby et al., 1994). In the present study also, only 13.51% isolates demonstrated the presence of hemolysin on blood agar. Tiwari et al. (2002) found only some of the 29 strains of Salmonellae to be weakly haemolytic.
Analysis for DNAse activity indicated 70.27% isolates to be positive. In an earlier study, 53 of 60 Salmonella strains were found to be DNAse positive and it was opined that the test may elserve as virulence and epidemiological marker (Chandra et al., 2007). Lysov and LuK (1991) found hospital originated strains to be highest DNAse producer.
DNase activity has been reported in S. Paratyphi B, S. Abortusequi and S. Weltevreden (Agarwal 2002; Sharma 2002; Kavitha et al., 2007). Our study also revealed that most of the serotypes isolated from diverse sources were positive for this activity in varying degree, indicating it to be wide spread phenomena. Role of DNAse in Salmonella pathogenesis is not clear but evidence from other bacteria indicates that DNAse has clear evidence in pathogenesis ( Sumby et al., 2005)
The mouse pathogenicity test by intra-peritoneal administration is an extensively used method for testing the virulence of Salmonella (Saxena et al., 2004) and considered superior over oral administration (Karim et al., 2008). In the present study, 100% mortality within 24 h was recorded for all the isolates tested indicating high virulence nature of the isolates. Variable mortality has been reported in earlier reports, which have been attributed to differential rate of clearance of pathogen (Saxena et al., 2004; Karim et al., 2008).
The study demonstrated that all the Salmonella isolates harboured inv and stn genes, whereas 94.59% of isolates had the presence of fimH and hilA genes. The isolates were also studied for cytotoxic, enterotoxic, Dnase and hemolysin activities which were found to be 72.97%, 51.35%, 70.27% and 13.51%, respectively. Besides, all the isolates were found to be highly pathogenic in mouse model. Among the different serotypes studied, Typhimurium and Paratyphi B var Java showed the presence of all the virulence determinants in varying degree. The 2 rough strains demonstrated least number of virulence determinants. They were devoid of fimH and hilA genes but harboured stn and invA genes. They were also positive for cytotoxic and enterotoxic activities. We could not trace any literature to compare our findings on rough strains. Tshiongwe and Labadi serotypes were also devoid of enterotoxigenic, DNAse and hemolytic activities. Other serotypes also lacked 1-2 virulence factors. In conclusion it can be said that Salmonella isolates studied in this report possessed diverse type of virulence determinants and were highly pathogenic. We also affirm that Salmonella pathogenecity is a complex mechanism and the organism uses different virulence determinants to inflict disease on the host.
REFERENCES
Abouzeed YM, Hariharan H, Poppe C and Kibenge FSB (2000). Characterization of Salmonella isolates from beef cattle, broiler chickens and human sources on Prince Edward Island. Comp Immun Microbiol Infect Dis. 23 :253–266.
http://dx.doi.org/10.1016/S0147-9571(99)00079-X
Agarwal M (2002). Molecular characterization of Salmonella Paratyphi var Java. M.V.Sc. Thesis, Deemed University, Indian Veterinary Research Institute, Izatnagar, India.
Agarwal RK (1997). Characterization of virulence factors of Aeromonas isolated from foods of animal origin. Ph.D. Thesis, Deemed University, IVRI, Izatnagar, Bareilly.
Agarwal RK, Bhilegaonkar KN, Singh DK, Kumar A and Rathore RS (2003). Laboratory manual for the isolation and identification of foodborne pathogen. Bareilly, Indian Veterinary Research Institute.
Ashkenazi S, Cleary TG, Murray BE, Wanger A and Pickering LK (1988). Quantitative analysis and partial characterization of cytotoxin production by Salmonella strains. Infect. Immun. 56 :3089–3094.
PMid:3182072 PMCid:PMC259706
Bajaj V, Hwang C and Lee CA. (1995). hilA is a novel ompR/toxR family member that activates expression of Salmonella Typhimurium invasion genes. Mol. Microbiol. 18 :715–727.
http://dx.doi.org/10.1111/j.1365-2958.1995.mmi_18040715.x
PMid:8817493
Baloda SB, Faris A, Krovacek K and Wadstrom T (1983). Cytotoxic enterotoxins and cytotoxic factors produced by S. enteritidis and S. typhimurium. Toxicon. 21 :785–790.
http://dx.doi.org/10.1016/0041-0101(83)90067-3
Baumler AJ, Tsolis RM and Heffron F (2000). Virulence mechanisms of Salmonella and their genetic basis. In: Salmonella in domestic animals. eds Wray, C., Wray, A. Wallingford, Oxford Shire, UK, CAB International, pp. 57–69.
Buzby JC, Roberts T Lin CTJ and MacDonald JM (1996). Bacterial Foodborne Disease: Medical Costs and Productivity Losses. Washington, DC. United States Department of Agriculture.
CDC. (2001). Diagnosis and management of foodborne illnesses: A primer for physicians. MMWR Recomm. Rep. 50 (1–69).
Chandra M, Singh, B.R., Shankar, H., Agarwal, M., Agarwal, R.K., Sharma, G. and Babu, N. (2007). Study on prevalence of Salmonella infection in goats. Small Rumin Res. 65: 24–30.
http://dx.doi.org/10.1016/j.smallrumres.2005.05.030
Chary P, Prasad R, Chopra AK and Peterson JW (1993). Location of the enterotoxin gene from Salmonella typhimurium and characterization of the gene products. FEMS Microbiol. Lett. 111:87–92.
http://dx.doi.org/10.1111/j.1574-6968.1993.tb06366.x
PMid:8395448
Chiu CH and Ou JT (1996). Rapid identification of Salmonella serovars in feces by specific detection of virulence genes, invA and spvC, by an enrichment broth culture–multiplex PCR combination assay. J. Clin. Microbiol. 34: 2619–2622.
PMid:8880536 PMCid:PMC229337
Clouthier SC, Muller KH, Doran JL, Collinson SK and Kay WW (1993). Characterization of three fimbrialgenes, sef ABC of Salmonella enteritidis. J. Bacteriol. 175 : 2523–2533.
PMid:8097515 PMCid:PMC204553
Cohen JI, Bartlett JA and Corey GR (1987). Extra-intestinal manifestations of Salmonella infections. Med (Baltimore). 66 (349–388).
http://dx.doi.org/10.1097/00005792-198709000-00003
Duncan MJ, Mann EL, Cohen MS, Ofek I, Sharon N and Abraham SN (2005). The distinct binding specificities exhibited by enterobacterial Type 1 - Fimbriae are determined by their fimbrial shafts. J. Biol. Chem. 280 :37707–37716.
http://dx.doi.org/10.1074/jbc.M501249200
PMid:16118220
Darwin KH and Miller VL (1999). Molecular basis of the interaction of Salmonella with the intestinal mucosa. Clin. Microbiol. Rev. 12 :405–428.
PMid:10398673 PMCid:PMC100246
Falkow F (1996). The Evolution of pathogenicity in Escherichia, Shigella, and Salmonella. In: Escherichia coli and Salmonella. ASM : 2723-2729.
Farmer JJ III (1995). Enterobacteriaceae: introduction and identification. In: Manual of clinical microbiology. eds Murray PR, Baron EJ, Pfaller MA, Tenover FC, Yolken RH. Washington DC, American Society for Microbiology Press. 438–449.
Frenzen P, Riggs T, Buzby J, Breuer T, Roberts T, Voetsch D, Reddy S and FoodNet Working Group. (1999). Salmonella cost estimate update using FoodNet data. Food Rev. 22 : 10–15.
Galan JE and Curtiss RV (1991). Distribution of the invA, B, C, and D genes of Salmonella Typhimurium among other Salmonella serovars: invA mutants of Salmonella Typhi are deficient for entry into mammalian cells. Infect. Immun. 59: 2901–2908. PMid:1879916 PMCid:PMC258111
Galan JE Ginocchio C. and Costeas P (1992). Molecular and functional characterization of the Salmonella invasion gene invA: homology of invA to members of a new protein family. J. Bacteriol. 174 :4338-4349.
PMid:1624429 PMCid:PMC206218
Giannella RA, Gots RE, Charney AN, Greenough WB and Formal SB (1975). Pathogenesis of Salmonella-mediated intestinal fluid secretion: activation of adenylate cyclase and inhibition by indomethacin. Gastroenterol. 69 :1238–1245.
Guo X, Chen J, Beuchat LR and Brackett RE (2000). PCR detection of Salmonella enterica serotype Montevideo in and on raw tomatoes using primers derived from hilA. Appl. Environ. Microbiol. 66 :5248-5252.
http://dx.doi.org/10.1128/AEM.66.12.5248-5252.2000
PMid:11097898 PMCid:PMC92452
Hu Q, Coburn B, Deng W, Li Y Shi, X Lan, Q Wang, Coombes BK and Finlay BB (2008). Salmonella enterica serovar Senftenberg human clinical isolates lacking SPI-1. J. Clin. Microbiol. 46 :1330–1336.
http://dx.doi.org/10.1128/JCM.01255-07
PMid:18272702 PMCid:PMC2292908
Houston CW, Koo FC and Peterson JW (1981). Characterization of Salmonella toxin released by mitomycin C-treated cells. Infect. Immun. 32 :916–926).
PMid:6788702 PMCid:PMC351529
Jiwa SFH (1981). Probing for enterotoxigenicity among the salmonellae: an evaluation of biological assays. J. Clin. Microbiol. 14 :463–472.
PMid:7309844 PMCid:PMC273970
Karim MR, Khan MSR, Kayesh MEH, Islam MR and Gani MO (2008). Isolation and characterization of sheep salmonellae in and around Bangladesh agricultural university campus. Bangl. J. Vet. Med. 6 : 75–78.
http://dx.doi.org/10.3329/bjvm.v6i1.1346
Kaura YK, Sharma VK and Chandiramani NK (1982). Enterotoxigenicity and invasiveness of Salmonella species. Antonie van Leeuwenhoek. 48: 273–283.
http://dx.doi.org/10.1007/BF00400387
PMid:6751222
Kavitha R, Chaturvedi VK, Pandey KD and Singh BR (2007). Heterogeneity among Salmonella enterica Weltevreden isolates of diverse host origin Indian J. Comp. Microbiol. Immunol. Infect. Dis. 27:10 – 13.
Ketyi I, Pacsa S, Emody L, Vertenyi A, Kovsis B and Kuch B (1979). Shigella dysenteriae 1-like cytotoxic enterotoxins produced by Salmonella strains. Acta Microbiol. Acad. Sci. Hung. 26 :217–223.
PMid:396765
Kuhn H, Tschape H and Rische H (1978). Einterotoxigenicity among salmonellae-a prospective analysis for a surveillance programme. Zentralbl. Bakteriol. Hyg. Abt. Orig. 240 : 171–183.
Libby SJ, Goebel W and Ludwig A (1994). A cytolysin encoded by Salmonella is required for survival within macrophages. Proc. Natl. Acad. Sci. USA. 91: 489–493.
http://dx.doi.org/10.1073/pnas.91.2.489
PMid:8290552 PMCid:PMC42974
Lysov PS and LuK R (1991). [DNAase activity, a method of differentiating hospital strains of Salmonella typhimurium. Lab. Delo. 11 :60–63. PMid:1722855
Makino S, KuraZono H, Chongsanguam M, Hyashi H, Cheun H, Suzuki S and Shirahata T (1999). Establishment of the PCR system specific to Salmonella spp. and its application for the inspection of food and fecal samples. J. Vet. Med. Sci. 61 : 1245–1247.
http://dx.doi.org/10.1292/jvms.61.1245
PMid:10593584
Menghistu HT (2010). Studies on molecular heterogeneity among Salmonella Gallinarum isolates of poultry origin. M.V.Sc. Thesis, Deemed University, IVRI, Izatnagar, Bareilly.
Mills DM, Bajaj V and Lee CA (1995). A 40 kb chromosomal fragment encoding Salmonella typhimurium invasion genes is absent from the corresponding region of the Escherichia coli K-12 chromosome. Mol. Microbiol. 15 : 749–759.
http://dx.doi.org/10.1111/j.1365-2958.1995.tb02382.x
PMid:7783645
Murugkar HV, Rahman H and Dutta PK (2003). Distribution of virulence genes in Salmonella serovars isolated from man and animals. Indian J Med. Res. 117 : 66–70.
PMid:12931840
O'Brien AD and Holmes RK (1996). Protein toxins of Escherichia coli and Salmonella, In: Escherichia coli and Salmonella: cellular and molecular biology ed Neidhardt FC, Curtiss III R, Ingraham JL, Lin ECC, Low KB, Magasanik B, Reznikoff WS, Riley M, Schaechter M and Umbarger HE, 2nd ed. Washington, D.C. American Society for Microbiology, pp. 2788–2802.
O'Brien AD, LaVeck GD, Thompson MR and Formal SB (1982). Production of Shigella dysenteriae type 1-like cytotoxin by Escherichia coli. J. Infect. Dis. 146 : 763–769.
http://dx.doi.org/10.1093/infdis/146.6.763
PMid:6754826
Okazaki N, Osawa R, Suzuki R, Nikkawa T and Whiley RA (2003). Novel observation of hot-cold-hot hemolysis exhibited by Group B Streptococci. J. Clin. Microbiol. 41: 877–879.
http://dx.doi.org/10.1128/JCM.41.2.877-879.2003
PMid:12574305 PMCid:PMC149725
Pathmanathan SG, Cardona-Castro N, Sanchez-Jimenez MM, Correa-Ochoa MM, Puthucheary SD and Thong KL (2003). Simple and rapid detection of Salmonella strains by direct PCR amplification of the hilA gene. J Med Microbiol. 52 (773–776).
PMid:12909653
Prager R, Fruth A and Tschape H (1995). Salmonella enterotoxin (stn) gene is prevalent among strains of Salmonella enterica, but not among Salmonella bongori and other Enterobacteriaceae. FEMS Immunol. Med. Microbiol. 12 :47–50.
http://dx.doi.org/10.1111/j.1574-695X.1995.tb00173.x
PMid:8580901
Rahman H (1999). Prevalence of enterotoxin gene (stn) among different serovars of Salmonella. Indian J. Med. Res. 110: 43–46.
PMid:10573653
Ray MK, Mitra A, Mukharjee B. and Chatterjee GC (1987) Study of Salmonella endotoxin on the changes in lipid-protein interactions of membranes using Arrhenius plots of cetylcholinesterase as a tool. J. Biosci. 11: 81–87.
http://dx.doi.org/10.1007/BF02704660
Sandefur PD and Peterson JW (1976). Isolation of skin permeability factors from culture filtrates of Salmonella Typhimurium. Infect. Immun. 14 : 671–679.
PMid:786890 PMCid:PMC420939
Saxena MA, Singh VP, Lackchura BD, Saxena A and Sharma B (2004). Detection of virulence in Indian isolates of Salmonella by polymerase chain reaction. Indian J. Biotechnol. 3: 37–40.
Sharma G (2002). Studies on double mutants (aroA-hirA deletion) of Salmonella Abortus eqi to evaluate long term production in guinea pigs against abortion and their in vitro and in vivo genetic stability. M.V.Sc. Thesis, Deemed University, Indian Veterinary Research Institute, Izatnagar, India.
Singh S, Agarwal RK, Tiwari SC and Kumar K (2009) Evaluation of a PCR assay for the detection of Salmonella from human clinical samples. Indian J. Comp. Microbiol. Immunol. Infect. Dis. 30 : 20–22.
Sumby P, Barbian KD, Gardner DJ, Whitney AR, Welty DM, Long RD, Bailey JR, Parnell MJ, Hoe NP, Adams GG, DeLeo FR and Musser JM (2005). Extracellular deoxyribonuclease made by group A Streptococcus assists pathogenesis by enhancing evasion of the innate immune response. Proc. Nat. Acad. Sci. 102 : 1679 –1684.
http://dx.doi.org/10.1073/pnas.0406641102
PMid:15668390 PMCid:PMC547841
Swamy SC, Barnhard HM, Lee MD and Dressen DW (1996). Virulence determinants invA and spvC in Salmonellae isolated from poultry products, waste water, and human sources. Appl. Environ. Microbiol. 62 : 3768–3771.
PMid:8837432 PMCid:PMC168184
Tiwari RP, Deol K, Rishi P and Grewal JS (2002). Factors affecting haemolysin production and Congo red binding in Salmonella enterica serovar Typhimurium DT 98. J. Med. Microbiol. 51: 503–509. PMid:12018658
Trafny EA, Kozłowska K and Szpakowska M (2006). A novel multiplex PCR assay for the detection of Salmonella enterica serovar Enteritidis in human faeces. Lett. Appl. Microbiol. 43: 673–679.
http://dx.doi.org/10.1111/j.1472-765X.2006.02007.x
PMid:17083715
Uppal MK, Rishi P and Tiwari RP (1998). Factors affecting hemolysin expression and its associated characteristics in Salmonella typhimurium and S. typhi. Ind. J. Med. Microbiol. 16: 169–174.
Van Asten AJAM and van Dijk JE (2005). Distribution of ''classic'' virulence factors among Salmonella spp. FEMS. Immunol Med Microbiol. 44 : 251–259.
http://dx.doi.org/10.1016/j.femsim.2005.02.002
PMid:15907446
Wallis TS, Starkey WG, Stephen J, Haddon SJ, Osborne MP and Candy DCA (1986). Enterotoxin production by S. Typhimurium strains of different virulence. J. Med. Microbiol. 21: 19–23.
http://dx.doi.org/10.1099/00222615-21-1-19
PMid:3512831
Watson PR, Galyov EE, Paulin SM, Jones PW and Wallis TS (1998). Mutation of invH, but not stn, reduces Salmonella-induced enteritis in cattle. Infect. Immun. 66: 1432–1438.
PMid:9529064 PMCid:PMC108071



