Advances in Animal and Veterinary Sciences
Research Article

Electron Microscopy and Immunohistochemistry using Indicators of Apoptosis and Proliferation on Guinea Pig Parathyroid Gland
Aya Hamdy Tantawy*, Ehab Mahmoud Elzoghby, Hossam Fouad Attia, Mahmoud Abdelghaffar Emam
Histology Department, Faculty of Veterinary Medicine, Benha University, Egypt.
Abstract | Maintenance of blood calcium level depended on the parathyroid hormone secretion that released from the parathyroid gland which important for bone mineralization. This work aimed to investigate the ultrastructure and immunohistochemical reaction of parathyroid gland of Guinea pig. The study was carried out on twenty five healthy male Guinea pigs, sixty days prenatal, one month postnatal and twelve months postnatal. Parathyroid gland consisted of fibrous capsule and cellular parenchyma which included chief and oxyphil cells. Ultrastructuraly, parathyroid gland contained several chief cells, fibroblast and blood capillaries. Chief cells included two types; granulated and vacuolated. The granulated cells contained several organelles and different sizes of secretory granules. The vacuolated cells contained few organelles and several vacuoles. The organelles involved mitochondria, Golgi complex, lysosome and different shapes of RER. Immunohistochemical studies were done using caspase3 and Bax as marker for apoptosis. Bax was responsible for activation of caspase3 via mitochondrial pathway of apoptosis. PCNA and Ki-67 were used as markers for cell proliferation. Ki-67 reacted during different stages of cell cycle and increased in S-phase. PCNA reacts mainly in S-phase of cell cycle, indicating condensation of DNA in the nucleus. Immunohistochemical staining with PCNA and Ki-67 shows negative and positive nuclear reaction at early life, then cells tend to increase their nuclear reaction with age. Caspase3 showed a faint cytoplasmic reaction at early life and that increased gradually with age. Bax showed a positive reaction in the cytoplasm and increased with age.
Keywords | Bax, Caspase 3, Guinea pig, Ki-67, Parathyroid gland and PCNA
Received | September 17, 2021; Accepted | October 28, 2021; Published | December 01, 2021
*Correspondence | Aya Hamdy Tantawy, Associate Lecturer of Histology, Faculty of Veterinary Medicine, Benha University, Egypt; Email: Aya.hamdy@fvtm.bu.edu.eg
Citation | Tantawy AH, Elzoghby EM, Attia HF, Emam MA (2022). Electron microscopy and immunohistochemistry using indicators of apoptosis and proliferation on guinea pig parathyroid gland. Adv. Anim. Vet. Sci. 10(1): 131-138.
DOI | http://dx.doi.org/10.17582/journal.aavs/2022/10.1.131.138
ISSN (Online) | 2307-8316; ISSN (Print) | 2309-3331
Copyright © 2022 Tantawy et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
INTRODUCTION
Parathyroid gland had special anatomy and physiology (Modarai et al., 2004). It had a great role in calcium homeostasis through production of parathyroid hormone (PTH). Serum calcium was played many physiological functions including blood coagulation and bone mineralization (Ramasamy, 2006). Parathyroid gland consisted of connective tissue capsule and cellular paranchyma which included chief cells, fibroblast and blood capillaries (Mini and Manju, 2017). There were two types of chief cells according to different stages of the secretory cycle; inactive and active chief cells (Chen et al., 2004).
The Guinea pig (Cavia porcellus) was one of the rodents that belonged to family Caviidae (Burnie, 2008). They were used in medical research, as a source of food in some countries, and as pet animal (Meredith and Redrobe, 2010; Gad, 2013). Studies (Nagpal et al., 1989; Ramayya et al., 2016) demonstrated that histological structure of parathyroid gland of Guinea pig was similar to that of mammals. Therefore, investigating the immunohistochemical reactions of parathyroid gland of Guinea pig will be a good indicator for other large animals.
Bax, caspase 3, PCNA and Ki-67 are common antibodies, which were used increasingly in immunohistochemical investigations. Bax and Caspases were among the most important antibodies used as markers for the apoptosis (Hadar et al., 2005). On the other hand, PCNA and Ki-67 were used as markers for cell proliferation, an indicator for parathyroid diseases (Carlson, 2010). Bax was a nuclear protein that penetrated the mitochondrial outer membrane to initiate apoptosis of cells. It was mainly present in the cystol of cell and little amount was located in the outer mitochondrial membrane and the endoplasmic reticulum (Dana et al., 2011). PCNA was synthesized during the G1 phase of the cell cycle, and reached its maximum amount during the S phase (Hofsta et al., 1995). Ki-67 was expressed in different stages of cell cycle. It increased in S- phase of cell cycle, but disappeard in resting phase (G0 phase) (Darzynkiewicz et al., 2015; Cuylen et al., 2016).
Although Guinea pig is an important laboratory animal, limited studies have been done on parathyroid gland of Guinea pig, so the current study was a trial to throw a light on the ultrastructure and immunohistochemical characteristics of parathyroid gland of Guinea pig.
MATERIALS AND METHODS
The present study was carried out on twenty five healthy Guinea pigs in the Department of Histology, Faculty of Veterinary Medicine, Benha University, Egypt.
Animals
A number of twenty one Guinea pigs were used for immunohistochemistry. According to Harkness et al. (2010) male Guinea pigs reached sexual maturity between 8 and 10 weeks. Therefore, ages of immature and mature animals were considered as follows:
| Age | Stage | Number |
| 60 day | Prenatal | 7 |
| 1 month | Immature postnatal | 7 |
| 12 months | Mature postnatal | 7 |
Also, four mature Guinea pigs (12 months) were used for electron microscope studies.
Sampling
Guinea pigs of different ages were obtained from the experimental animal house at the Zoo, Giza Governorate, Egypt. All animals were examined before processing to be free from any abnormal disease. The animals were euthanized in compliance with recommendations from the animal care committee of the Faculty of Veterinary Medicine, Benha University (approval number: BUFVTM 02-08-21). Opening the neck region is the way to obtain the thyroid and parathyroid gland from the middle third of tracheal rings. Then parathyroid gland was separated from thyroid gland.
Electron microscopy
Specimens preparations for electron microscopy investigation were done according to procedures outlined by Bozzola and Russsell (1999).
Transmission electron microscopy
1mm³ specimens were used for transmission electron microscope study. Specimens were fixed in 2.5% glutaraldehyde in 0.1 M sodium cacodylate buffer, and post fixated for 2 hours in 1% osmium tetroxide buffered to pH 7.4 at 4°C. The fragments were dehydrated in a graded series of ethyl alcohol and embedded in epoxy resin. Semithin sections 0.5µm in thickness were used for histological observations after methylene blue staining. The ultra-thin sections were prepared with ultra-cut microtome, stained with uranyl acetate and lead citrate and examined by a Joel JEM-100S transmission electron microscope (25KV) at the faculty of science, Alexandria University. Representive fields of each specimen were photographed for detailed image analysis.
Immunohistochemistry
Paraffin sections of 5 micrometer from the parathyroid gland were collected on positive charged microscope slides. Sections were deparaffinezed in xylene, rehydrated sequentially in absolute ethanol, 95% ethanol, 70% ethanol, distilled water and rinsed in phosphate buffered saline.
Antigen retrieval was done by heating the tissue sections in 10mM citrate buffer, PH 6.0 for 40 minutes followed by cooling at room temperature for 20 minutes. For reducing peroxidase activity, sections were treated with 3% hydrogen peroxide in methanol for 15 minutes. To block non-specific stain, sections were incubated with 10% normal goat serum for 30 minutes at room temperature.
Sections were incubated with rabbit anti-human Bax polyclonal antibodies (catalog number E17990, Spring Bioscience, Pleasanton, California, USA), at dilution 1:200 at room temperature for 30 minutes. Then the sections were treated with goat anti-rabbit secondary antibody for 30 minutes at room temperature.
Sections were incubated with rabbit polyclonal caspase3 antibodies (catalog number C9598, Sigma Aldrich, Street Louis, Missouri, USA), at dilution 1:1000 at room temperature for 30 minutes. Then the sections were treated with human anti-rabbit secondary antibody for 30 minutes at room temperature.
Sections were incubated with mouse monoclonal PCNA antibodies (catalog number P8825, Sigma Aldrich, Street Louis, Missouri, USA), at dilution 1:500 at room temperature for 30 minutes. Then the sections were treated with mouse secondary antibody for 30 minutes at room temperature.
Sections were incubated with mouse monoclonal Ki-67 antibodies (catalog number API 3156 AA, Sigma Aldrich, Street Louis, Missouri, USA), at dilution between 1:50 to 1: 500 at room temperature for 30 minutes. Then the sections were treated with mouse secondary antibody for 10 minutes at room temperature. Visualization was done as outlined by Kumar and Rudbeck (2009).
RESULTS AND DISCUSSION
Transmission electron microscope
Electron microscope is done on mature parathyroid glands of twelve months Guinea pig. A semi thin section of one year parathyroid gland consists of fibrous capsule and numerous chief cells (Figure 1A). Chief cells are granulated and vacuolated cells between blood capillaries (Figure 1B).
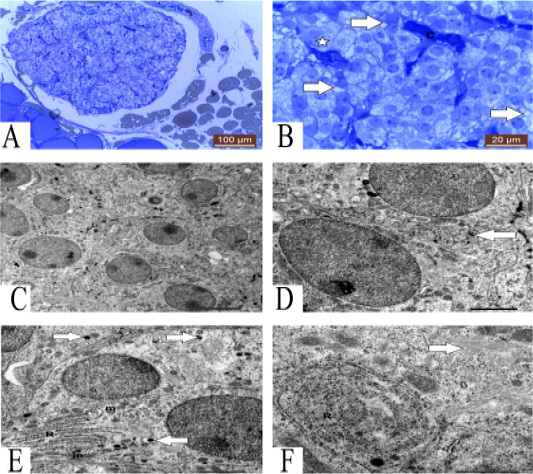
Figure 1: A: Photomicrograph showing semithin section of parathyroid gland of one year guinea pig. Methylene blue (100µm). B: Photomicrograph showing semithin section of parathyroid gland showing granulated (star), vacuolated (arrows) chief cells and blood capillaries (C). Methylene blue (20µm). C: TEM of mature parathyroid gland showing several chief cells with different organelles 5 μm. D: TEM of mature parathyroid gland showing tortuous plasma membrane (arrow) of chief cells 2 μm. E: TEM of chief cells contain RER (R), mitochondria (m) and secretory granules (arrows) 2 μm. F: TEM of chief cells contain concentric RER (R) and Golgi complex (arrow) 1 μm.
Parathyroid gland contains several chief cells with different organelles (Figure 1C). Plasma membrane of chief cells has straight course, but sometimes plasma membrane of some cells has tortuous course (Figure 1D).
Chief cells contain several organelles, lysosome, rough endoplasmic reticulum (RER) with flattened cisternae, mitochondria and secretory granules (Figure 1E). Golgi complex and characteristic concentric cisternae of RER also present (Figure 1F). Mitochondria have several forms round and oval mitochondria (Figure 2A). Some chief cells have undulating nucleus with condensation of chromatin at certain areas in the nucleus, large secretory granules and some vacuoles (Figure 2B). Also, other chief cells contain several chromatin islands, secretory granules and RER with concentric cisternae (Figure 2C).
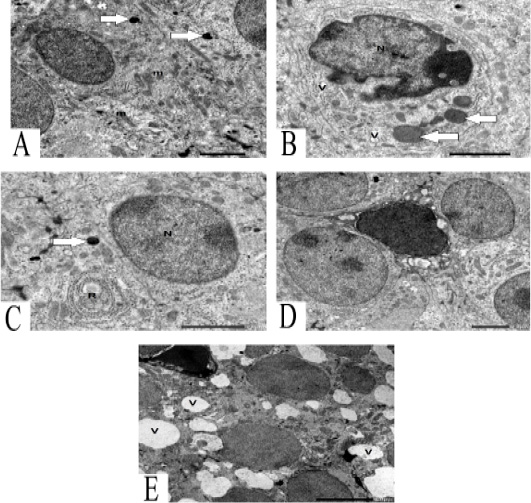
Figure 2: A: TEM of chief cells contains round and oval mitochondria (m) and lysosome (arrows) 2 μm. B: TEM of chief cells contain undulating nucleus (N) with condensation of chromatin, large secretory granules (arrows) and some vacuoles (V) 2 μm. C: TEM of chief cells contains nucleus (N) with chromatin islands, secretory granules (arrow) and characteristic RER (R) 2 μm. D: TEM of vacuolated chief cells between granulated chief cells. 2μm. E: TEM of vacuolated chief cells with dark nucleus and several vacuoles (V) .5 μm.
The parathyroid gland contains vacuolated chief cells between granulated chief cells (Figure 2D). Vacuolated chief cells contain dark heterochromatic nucleus and several vacuoles (Figure 2E).
Immunohistochemistry
PCNA and Ki-67
Parathyroid gland of 60 embryonic days (ED) Guinea pig show negative and positive immunoreactivity to PCNA. Some cells show negative nuclear reaction. Positive reactive cells are two types; some cells show strong nuclear reaction indicating condensation of DNA strands and more viable cell. On the other hand, some cells show weak nuclear reaction representing low condensation of DNA strands (Figure 3A).
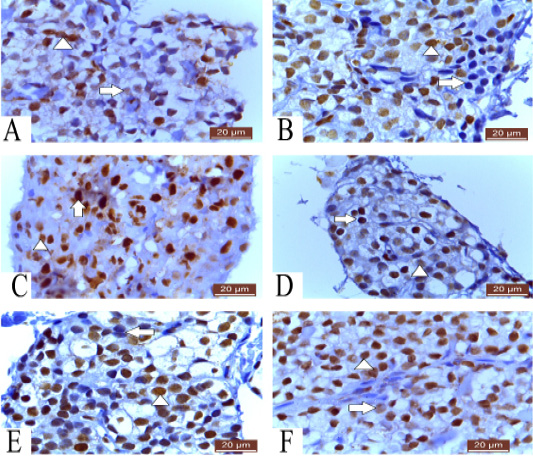
Figure 3: A: Photomicrograph of parathyroid gland of 60 ED showing negative nuclear reaction of few cells, strong nuclear reaction of some cells (arrowhead) and weak reaction of others (arrow) PCNA (20µm). B: Photomicrograph of parathyroid gland of 60 ED Guinea pig showing few cells give negative nuclear reaction (arrow) and most of cells show positive nuclear reaction (arrowhead) Ki-67 (20µm). C: Photomicrograph of parathyroid gland of 1 month showing strong nuclear reaction of some cells (arrow) and weak reaction of others (arrowhead) PCNA (20µm). D: Photomicrograph of parathyroid gland of 1 month Guinea pig showing strong (arrow) and weak (arrowhead) positive nuclear reaction Ki-67 (20µm). E: Photomicrograph of parathyroid gland of 12 months Guinea pig showing few negative (arrow) and most positive nuclear reaction (arrowhead) Ki-67 (20µm). F: Photomicrograph of parathyroid gland of 12 months showing strong nuclear reaction of most of cells (arrow head) and weak reaction of few cells (Arrow) PCNA (20µm).
Parathyroid cells of 60 ED Guinea pig show negative and positive nuclear reaction. Some cells react negatively with ki-67. Most of cells are positively reacted (Figure 3B).
Parathyroid gland of 1 month Guinea pig showing variable forms of cell viability, some cells show dense nuclear reaction to PCNA indicating more viable cell which present at late S- phase of cell cycle. On the other hand, some cells show weak nuclear reaction representing cell present in early S- phase of cell cycle (Figure 3C).
Parathyroid gland of 1 month Guinea pig show weak and strong positive nuclear immunoreactivity to ki-67 (Figure 3D).
Also, parathyroid cells of 12 months Guinea pig show both negative and positive nuclear reaction, few numbers of cells show negative reaction and most of cells are positively reacted to ki-67 (Figure 3E). However, parathyroid cells of 12 months Guinea pig show strong nuclear reaction and few numbers of cells show faint reaction to PCNA (Figure 3F).
Caspase 3 and Bax
Parathyroid cells of 60 ED Guinea pig show very weak cytoplasmic reaction to caspase 3 (Figure 4A). While, parathyroid gland of 60 ED Guinea pig show negative and positive immunoreactivity to Bax. Some cells show negative cytoplasmic reaction. Few numbers of cells show positive cytoplasmic reaction (Figure 4B).
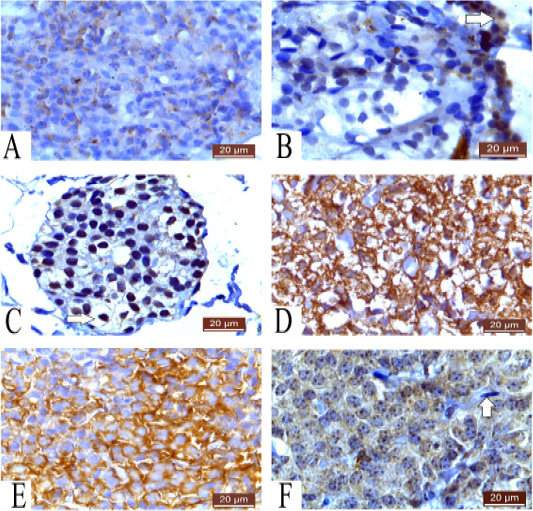
Figure 4: A: Photomicrograph of parathyroid gland of 60 ED showing very weak cytoplasmic reaction Caspase 3 (20µm). B: Photomicrograph of parathyroid gland of 60 ED Guinea pig showing negative cytoplasmic reaction of most of cells and positive cytoplasmic reaction of some cells (arrow) Bax (20µm). C: Photomicrograph of parathyroid gland of 1 month Guinea pig showing positive reaction and negative cytoplasmic reaction (arrow) Bax (20µm). D: Photomicrograph of parathyroid gland of 1 month showing positive cytoplasmic reaction of cells Caspase3 (20µm). E: Photomicrograph of parathyroid gland of 12 months showing strong cytoplasmic reaction of most of cells Caspase 3 (20µm). F: Photomicrograph of parathyroid gland of 12 months Guinea pig showing positive cytoplasmic reaction and negative fibroblast (arrow) Bax (20µm).
Parathyroid gland of 1 month Guinea pig showing positive reaction in large number of cells, while few number of cells show negative cytoplasmic reaction to Bax (Figure 4C).
Parathyroid gland of 1 month Guinea pig show positive immunoreactivity of caspase 3 in the cytoplasm of cells and few cells show faint reaction (Figure 4D).
However, parathyroid cells of 12 months Guinea pig show strong cytoplasmic reaction to Caspase 3 (Figure 4E) and most parathyroid cells of 12 months Guinea pig show strong cytoplasmic reaction and fibroblast show negative reaction to Bax (Figure 4F).
Parathyroid gland produces PTH which was responsible for calcium hemostasis (Ramasamy, 2006). The PTH was firstly synthetized as Pre-proparathyroid hormone on ribosomes of RER, composed of 115 amino acids and rapidly converted to proparathyroid hormone. The intermediate precursor was composed of 90 amino acids and moves within channels of the endoplasmic reticulum to the Golgi apparatus. Enzymes within membranes of the Golgi apparatus were responsible for formation of active PTH that aggregated in the Golgi apparatus of chief cells. Finally, C- and N-terminal were formed in the peripheral circulation as end products of PTH (Cohn and MacGregor, 1981).
Guinea pig parathyroid gland consisted of fibrous capsule and cellular parenchyma which contained numerous blood capillaries. This structure is similar to that of rat, mice and hamster (Basha and Wood, 1990). Commonly, chief and oxyphil cells were the main cellular components of parathyroid gland. However, at young ages, parathyroid gland of Guinea pigs only contained chief cells which appeared as irregular clusters, cords and follicles around blood capillaries. These findings were in line with those reported for most rodents (Nakanishi et al., 2004) as well as human (Chen et al., 2013; Mini and Manju, 2017).
Chief cells were polyhedral cells with round or oval nuclei. The cytoplasm of these cells appeared in different forms either granular or vacuolated and sometimes empty cells according to secretory activity. These results were in line with those reported for rat and mice (Enemali et al., 2017; Birgit et al., 1996) and human (Mini and Manju, 2017). The dark chief cells were polygonal shape with oval or round nuclei and dark cytoplasm. The cytoplasm was dark owing to presence of several organelles and numerous secretory granules that indicated active protein synthesis. Increased number of rough endoplasmic reticulum and Golgi apparatus were important for synthesis of PTH (Utsumi et al., 1999). On the other hand, the light chief cells were characterized by light cytoplasm due to less cell organelles and more vacuolated cytoplasm. Similar results were recorded in camel (Al-Ramadan et al., 2016) and human (Chen et al., 2013).
Chief cells of Guinea pig contained several secretory granules, small and large similar to mouse, rat and hamster (Isono et al., 1985; Utsumi et al., 2014). However, these results disagreed with rat chief cells contained rare secretory granules as the secretory activity occurs in parathyroid glands of embryonal and fetal rat only (Isono et al., 1990). Some chief cells contained vacuoles which were important in exocytosis of secretory granules. This result was similar to mouse chief cells. Also, the plasma membrane of adjacent chief cells was of straight course and some cells had tortoise plasma membrane. This was similar to chief cells of mouse (Ishizaki et al., 1989).
Apoptosis or programmed cell death was a regulatory process for maintenance of hemostasis during normal growth and development. Apoptosis was a line of defense against infectious and immunological diseases (Milligan and Schwartz, 1997; Thornberry and Lazebnik, 1998). Parathyroid diseases were detected by using numerous markers of apoptosis like caspase 3 and Bax (Hadar et al., 2005).
Bax had an important role in activation of caspase 3 through the intrinsic pathway of apoptosis. This pathway occurred when Bax protein initiated releasing of cytochrome c protein from the mitochondria. Cytochrome c was bonded with apoptotic protease activating factor 1 and ATP, then bonded to pro-caspase 9. This protein complex cleaved to active form of caspase 9 that cleaved and activated to caspase 3 (Uren et al., 2017).
Successful apoptosis depended on inducers, caspase enzymes, related genes and signaling pathways (Parrish et al., 2013). Also, there were 14 types of caspases in humans that activated by different death stimuli in different cell types. Caspase-3 had a great role in apoptosis as it was responsible for chromatin condensation, DNA fragmentation (Porter and Jänicke, 1999) and degradation of nuclear polymerase (Payam and Reza, 2015). Caspase-3 could be activated in the cell both by extrinsic and intrinsic pathways of apoptosis (Ghavami et al., 2009).
Parathyroid cells were demonstrated weak cytoplasmic reaction to caspase 3 and Bax at the young Guinea pig and became stronger with age representing more apoptotic cells (Renehan et al., 2001).
Cell proliferation was a process where the cell undergoes cell division and cell growth for production of two daughter cells and maintenance of cellular homeostasis. In case of abnormal cell proliferation, cancer might occur (Van Diest et al., 1998; Grewal and Edgar, 2003).
PCNA was a clamp of DNA that work on DNA polymerase delta in eukaryotic cells and was important for DNA replication and DNA repair (Moldovan et al., 2007). It was expressed in the nucleus of cells during cell cycle to give an indication for DNA condensation in the nucleus and viability of cells (Leonardi et al., 1992). It had an important role in differentiation between early, mid and late S phase of the cell cycle through imaging of the nuclear distribution of PCNA (Schönenberger et al., 2015).
Cells of parathyroid gland of Guinea pig showed negative and positive nuclear immunoreactivity to PCNA. There were strong and weak positive reaction, strong reaction indicating more synthesis of DNA and more viable cell and weak reaction representing low condensation of DNA strands. The positive nuclear reaction was increase with aging. These was similar to that is demonstrated in human and mice (Leonardi et al., 1992).
Ki-67 was a nuclear protein, expresses in different stages of cell cycle, G1, S, G2 and mitosis. It was increased in S- phase of cell cycle (Booth and Earnshaw, 2017). Ki-67 had a great role in cell proliferation; during interphase, it was important for normal cellular distribution of heterochromatin antigens and for the attachment of heterochromatin to the nucleolus, while during mitosis, it was required for formation of the perichromosomal layer which coated the condensed chromosomes. Also, Ki-67 was responsible for preventing the sticking of mitotic chromosomes (Xiaoming and Paul, 2018). The perichromosomal layer was a proteineous sheath located at the outer surfaces of individual chromosomes, represented one-third of the protein mass of mitotic chromosomes (Booth and Earnshaw, 2017). Ki-67 was one of the earliest proteins associated with perichromosomal layer and stayed on it till telophase (Van Hooser et al., 2005).
Parathyroid cells in early stages of cell cycle were showed weak nuclear expression to Ki-67, which increased with re-entering the cell cycle. With aging, cells reacted positively in a gradual manner that indicated increasing cell proliferation (Sobecki et al., 2017).
CONCLUSIONS AND RECOMMENDATIONS
In the current work, the electron microscope study indicated that parathyroid gland contained vacuolated chief cells between granulated cells. The chief cells were characterized by the presence of concentric cisternae of RER. There were several forms of secretory granules (small and large). Using Bax and caspase 3 as markers for apoptosis, parathyroid cells showed weak cytoplasmic reaction at early life then increased with aging. Parathyroid cells showed negative and positive nuclear reaction with PCNA and Ki-67 that increased with age. Therefore, with aging, cells reacted positively in a gradual manner that indicated increasing cell proliferation. Further studies are recommended for deep understanding the ultrastructure and immunohistochemical studies of parathyroid gland of Guinea pig.
ACKNOWLEDGMENTS
The authors wish to express their gratitude to Faculty of Veterinary Medicine, Benha University, Egypt for supporting this work.
Novelty Statement
Ultrastructural and immunohistochemical investigations were done on parathyroid gland of Guinea pig at different developmental stages (prenatal and postnatal). Using TEM, the concentric cisternae of RER were the most predominant feature of chief cells. Markers of apoptosis and proliferation showed weak reaction at early life stage and increased gradually with age.
Author’s Contribution
EME, HFA & MAE: Idea and design. AHT: Sample collection and perpetration, electron microscopy and immunohistochemical examinations. AHT & MAE: Write the manuscript. EME & HFA: Revision.
Conflict of interest
The authors have declared no conflict of interest.
REFERENCES



