Journal of Infection and Molecular Biology
Research Article
Journal of Infection and Molecular Biology. 1 (3): 41 – 45Segregation of Plasma Cells in Lymphoid Organs by Various Routes of Vaccination against Newcastle Disease in Broiler Chickens
Tasra Bibi1*, Asim Aslam1, Shafqat Fatima Rehmani2, Arfat Yousaf Shaheen3, Nadia Mukhtar3, Abdul Wajid4, Saira Jabeen3, Suliman Qadir Afridi3, Hassan Anjum Shahid5
- Department of Pathology, University of Veterinary and Animal Sciences Lahore 54000
- Quality Operation Laboratory (QOL), University of Veterinary and Animal Sciences Lahore 54000
- Department of Microbiology, University of Veterinary and Animal Sciences Lahore 54000
- Institute of Biochemistry and Biotechnology (IBBt), University of Veterinary and Animal Sciences Lahore 54000
- King Edward Medical University, Lahore, Pakistan
*Corresponding author: tasraanjum@gmail.com
ARTICLE CITATION:
Bibi T, Aslam A, Rehmani SF, Shaheen AY, Mukhtar N, Jabeen S, Wajid A, Afridi S, Shahid HA (2013). Segregation of plasma cells in lymphoid organs by various routes of vaccination against newcastle disease in broiler chickens. J. Inf. Mol. Biol. 1 (3): 41 – 45.
Received: 2013–03–07, Revised: 2013–07–03, Accepted: 2013–07–05
The electronic version of this article is the complete one and can be found online at
(
http://nexusacademicpublishers.com/table_contents_detail/2/63/html
)
which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited
ABSTRACT
One hundred ninety five (195) chicks were used to collect three different lymphoid organs i.e Thymus, Bursa of Fabricious and Harderian gland after vaccinating with Newcastle disease virus to observe the histopathological changes. The chicks were divided into three groups and vaccinated via coarse spry, eye drop and drinking water at the age of five days and then boasted at twenty six days of age. The histopathological changes show high number of Plasma cells in these lymphoid organs. The segregation of plasma cells was significantly higher in Harderian gland using NDV vaccine by eye drop method. All of the three routes of immunization provided comparable protection against the challenged infection at 6 weeks of age. However, the coarse spray method of vaccination provided significantly high level of circulating antibodies as compared to ocular and drinking water vaccination.
INTRODUCTION
Newcastle disease (ND) is an infectious, highly contagious and widespread disease of avian species. It has a considerable economic impact on poultry industry, especially chickens. Since its recognition in 1926 in Indonesia and Newcastle–upon–Tyne, England also occurred in Ranikhet, India in 1927, ND is regarded as being endemic in many countries (Bwala et al., 2009). The morbidity and mortality of susceptible birds is 100% in the severe form of the disease and unvaccinated birds are more prone to the disease (Alexander, 2003; Manchang et al., 2004). The poultry industry loses millions of dollars annually from mortality due to ND. Its importance is mainly due to the resulting economic losses that occur upon infection with virulent strains (Susta et al., 2011; Diel et al., 2012).
Immune responses of lymphoid organs to ND vaccine are influenced by routes (ocular, injection, drinking water, aerosol or nasal) of vaccination. Vaccines given through mucosalsurfaces (oral, eye drop and nasal) generate mucosal immune responses so they are more effective routes of vaccination for viruses entering in the body through mucosal surfaces, such as NDV (Salam, 2003). As the vaccinal strain is deposited directly onto respiratory mucosa, the spray route is suitable for respiratory type vaccines. Coarse spray is preferred vaccination method as it is proposed to deliver large droplets of vaccine to the upper respiratory tract and eye. In addition the moistened feathers of birds stimulating preening which further increase the chance of vaccine uptake (Breytenbach, 2005).
Giambrone (1985) reported that birds vaccinated with a live vaccine by coarse spray have highest NDV HI titers; resistance to challenge is also greatest. Eye drop or oral administration of NDV vaccines were also proved to be effective in eliciting mucosal immunity mediated by IgA antibodies as reported by (Jayawardane and Spradbrow, 1995; Parry and Aitken, 1973). Inactivated vaccines do not induce local immunity in the respiratory and digestive tracts; however, immunity is established rather slowly. Also the killed vaccines are expensive and difficult to administer than live vaccines (Van Eck, 1987).
The humoral and cell–mediated immunity acquired by vaccination play vital role for protecting the birds against field infection (Chandraseker et al., 1989). Live vaccines provide better protection as they provide instant protection to field virus challenge on later exposure. A vaccine that induces an immune response by uplifting mucosal immunity is a valuable way of targeting the respiratory pathogens before infection occurs (Brandtzaeg, 2004; Holmgren, 2005). Mucosal vaccination, in contrast to parenteral vaccination can produce both systemic and mucosal immunity by secreting both IgA and IgG immunoglobulins whereas parenteral vaccines only induce serum IgG antibodies (Yuki, 2003). This study was carried out for comparing the immune status of the broiler chickens and the morphological changes in lymphoid organs and its role, if any, to elicit the antibody response and to assess the efficacy of various routes of vaccination.
MATERIALS AND METHODS
Experimental Chicks
Day– old broiler chicks (n= 195) were procured from local commercial hatchery. They were brooded for 5 days in a semi specified pathogen free area at the University of Veterinary and Animal Sciences, Lahore.
Vaccination of Experimental Chicks
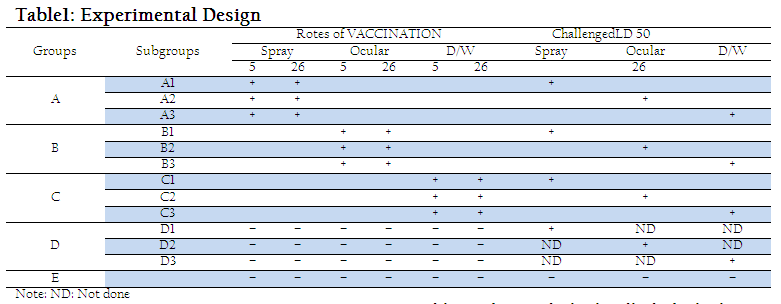
A commercially manufactured LaSota virus vaccine was purchased from the local market and administered via spray, eye droppings and drinking water route (Table–1).
Challenge Virus
A virulent field isolate of velogenic NDV (vNDV) previously characterized as velogenic by the research workers (unpublished data) in the (Quality Operation Laboratory), University of Veterinary and Animal Sciences, Lahore was selected. The biological titer of the virus was 105.5 LD50 and the challenge dose was 0.1 ml (100 × LD50) per bird.
On day 26, all experimental birds were challenged with velogenic NDV at dose rate of 0.1ml/ bird through Eye drop, drinking water and spray as shown in the Table 1.
Experimental Design
The experiment lasted for 42 days. On day 5, birds were individually identified with numbered wing tags after being randomly divided into four treatment groups i.e. A, B, C, D and one control group E containing 45 birds each. Then chicks of former four groups were subdivided into A1, A2,A3, B1, B2, B3,C1, C2,C3,D1,D2,and D3 containing 15 birds each on day 26. The experiment design is given in Table 1.
Collection of Samples
Blood samples were collected from 5 birds of each subgroup selected randomly on days, 5, 12, 19, 26, 33, 40 and serum samples were separated to assess the antibody titers against NDV. Lymphoid organs (Harderian gland, Bursa of Fabricius and thymus) from 5 randomly selected birds of each subgroup were collected on days 26, 33, 40 for histo–pathological studies.
Antibody titration against NDV
After collection, serum samples were stored at –40 C till used. Antibody titer against NDV in serum was determined by haemagglutination inhibition (HI) test as described by (Allan
and Gough, 1974).
Histopathology of Lymphoid Organs
Harderian Gland, bursa of Fabricius and thymus were collected and kept in 10% Neutral Buffered Formalin (NBF) in pre labeled containers. They were allowed to fix for 48 hours and were processed for histopathological studies (Nakamura et al., 2008; Bwala et al., 2012; Zakeri and Kashefi, 2012).
Plasma cell counting in Harderian gland
Plasma cells were counted in separate Harderian gland fields (200 x). Plasma cells numbers were expressed as Plasma cells per mm2 (Salam, 2003).
Post–challenge Mortality
Post–challenge mortality was recorded in all experimental birds. The symptoms and lesions were recorded by postmortem examination of the dead birds.
Statistical analysis
The data collected was statistically analyzed by Analysis of Variance and least significant difference (LSD) using SPSS 16.0 software , significance was accepted at P<0.05.
RESULTS AND DISCUSSION
Newcastle disease continues to be a major threat to the poultry industry despite the wide spread use of various vaccines and route of vaccination. One way of controlling this problem is to improve the efficacy of vaccines by experimenting different routes of immunization. In the present study, three routes (spray, ocular and drinking water) of vaccination against ND were used on 5 and 26 days of age. Geometric mean titers (GMT) of different groups at different days are shown in table 2. GMT of groups A, B and C showed a rise in antibody titer from 12th day of age, which reached the peak level at day 26. There was higher rise in antibody titer in group A and B as compared to group C as gastric secretions provided a non–specific barrier against invaders and destroyed them. Thus, some of the vaccinal virus given through oral route got denatured resulting in reduced antibody titer in group C (Tizard, 1996). Decrease in HI antibody titer was observed in subgroups A1, A2, A3, B1, B2, B3, C1, C2 and C3 and sharp rise in antibody titer in sub groups D1, D2 and D3 on 33th day of experiment (7 day post challenge), as shown in Table 2. These observations are in coordination with (Tizard, 1996) who described a decrease in antibody titer due to neutralization of virus with circulating antibodies and rise in antibody titer due to activation of immune system against challenge due to which rise in antibody titer noted in serum.
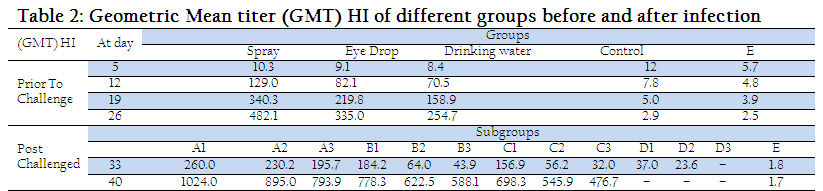
A sharp rise in antibody titer was also observed in subgroups A1, A2, A3, B1, B2, B3, C1, C2 and C3 on day 40 (14 days post challenge). Similarly (Manzoor, 1999) observed that in vaccinated birds challenged virus was neutralized by circulating antibodies and the immune system was boosted up resulting in increase in antibody titer. However, no histopathological changes were shown in parts of Harderian gland, bursa of fabricius and thymus of groups A, B, C, D and E on day 26, Table 3.
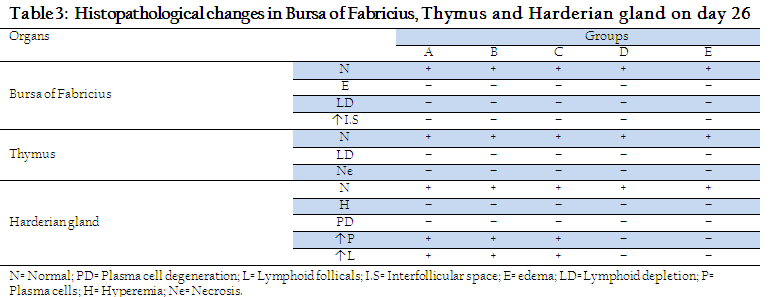
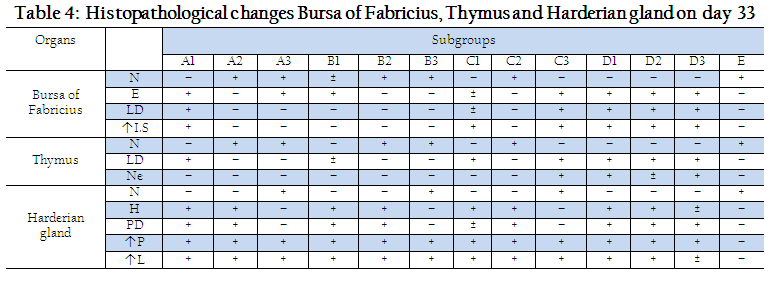
Histopathological changes in Harderian gland, bursa of fabricius and thymus of different groups at day 33 and 40 are shown in Tables 4 and 5. On day 33, no histopathological lesions were noted in Harderian Gland of Subgroups A3, B3, C3 and D3 and E. On day 33, no histopathological lesions were seen in bursa of fabricius and thymus of subgroups B2, B3, C2 and E, while severe lesions were seen in subgroups D1, D2 and D3 (Table 4). Histopathological lesions in Harderian Gland, Interfollicular edema and necrosis of lymphoid follicles were seen in bursa of Fabricius while thymus showed necrotic centers and less dense population of lymphocytes in subgroups A1, A2 and A3. A few birds of subgroup C3 also showed mild degree of same lesions. Our results are similar to previous reports (Salam, 2003; Giambrone, 1985) Who noted necrosis and degeneration in bursa of fabricius and thymus after NDV challenge.
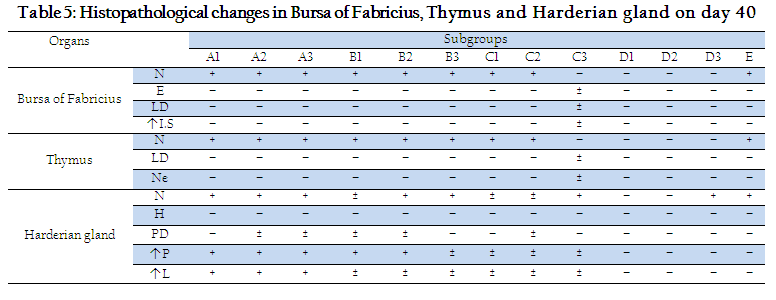
Mean plasma cell counts in sections of Harderian gland of different groups at different days are presented in Table–6. On day 26, significantly (P<0.05) higher number of Plasma cells were seen in stroma of Harderian gland in groups A, B and C as compared to D and E. These results are in line with the findings of (Jayawardane and spradbrow, 1995; Giambrone, 1985) who studied significant increase in plasma cells in sections of Harderian gland after vaccinations. Plasma cell count in group A was significantly (P<0.05) higher than that of group B and C. Hyperemia and vascularization along with increased number of plasma cells were seen in subgroups A1, A2, B1, B2, C1, C2, D1 and D2. Our findings are supported by those of (Salam, 2003). Increase in plasma cell number was also observed in orally challenged birds (Subgroups A3, B3, C3 and D3) but this increase was not as significant as that of coarse spray challenged birds (Table 6).


No mortality at twenty six (26) days post challenge of different subgroups A1, A2, A3 and B1 was recorded. This shows that the spray method of vaccination not only provides good protection against field challenge but also no pathological changes observed in the lymphoid organs.
ACKNOWLEDGMENTS
The authors acknowledged the cooperation of the Department of Pathology and Quality Operation Laboratory of the University of Veterinary and Animal Sciences, during the trial.
REFERENCES
Allan WH and Gough RE. (1974). A standard haemagglutination inhibition test for Newcastle disease. I. A comparison of macro and micro methods. Vet. Rec. 95: 120-123.
http://dx.doi.org/10.1136/vr.95.6.120
http://dx.doi.org/10.1136/vr.95.7.147
PMid:4446306
Alexander D. (2003). Newcastle disease, other avian paramyxoviruses, and pneumovirus infections.In: Brandtzaeg P and Pabst R. (2004). Let's go mucosal: communication on slippery ground. Trends Immunol. 25: 570–577.
Breytenbach, JH and Intervet International BV. (2005). Correct and effective vaccination of poultry. International Poultry Production.12: 17-21.
Bwala, DG, Clift S, Duncan NM, Bisschop SP, Oludayo FF. (2012). Determination of the distribution of lentogenic vaccine and virulent Newcastle disease virus antigen in the oviduct of SPF and commercial hen using immunohistochemistry. Res Vet Sci. 93: 520-528.
http://dx.doi.org/10.1016/j.rvsc.2011.06.023
PMid:21774952
Bwala DG, Abolnik C, van Wyk A, Cornelius E, Bisschop SP. (2009). Efficacy of a genotype 2 Newcastle disease vaccine (Avinew) against challenge with highly virulent genotypes 5d and 3d. J S Afr Vet Assoc. 80: 174-178
http://dx.doi.org/10.4102/jsava.v80i3.197
PMid:20169751
Chandraseker S, Venkatesan RA, Padmanaban VD, Masiilamony PR. (1989). Nature of protective immunity responses in chicken against Ranikhet disease. Ind. Vet. J. 66: 801-806.
Diel DG, Susta L, Cardenas Garcia S, Killian ML, Brown CC, Miller PJ and Afonso CL. (2012). Complete genome and clinicopathological characterization of a virulent Newcastle disease virus isolate from South America. J Clin Microbiol. 50: 378-387.
http://dx.doi.org/10.1128/JCM.06018-11
PMid:22135263 PMCid:PMC3264182
Giambrone JJ. (1985). Laboratory evaluation of Newcastle disease vaccination programs for broiler chickens. Avian Dis. 29: 479-487.
http://dx.doi.org/10.2307/1590509
PMid:4026739
Holmgren J and Czerkinsky C. (2005). Mucosal immunity and vaccines. Nat. Med. 11 (4, Suppl) S45–S53.
http://dx.doi.org/10.1038/nm1213
PMid:15812489
Jayawardane GWL, Spradbrow PB. (1995). Mucosal immunity in chickens vaccinated with the V4 strain of Newcastle disease virus. Vet. Microbiol. 46: 69-77.
http://dx.doi.org/10.1016/0378-1135(95)00073-J
http://dx.doi.org/10.1016/0378-1135(95)00068-L
Kim SH, Xiao S, Shive H, Collins PL, Samal SK. (2012). Replication, neurotropism, and pathogenicity of avian paramyxovirus serotypes 1-9 in chickens and ducks. PLoS One. 7: e34927.
http://dx.doi.org/10.1371/journal.pone.0034927
PMid:22558104 PMCid:PMC3340391
Majid-Zadeh A, Kita J, Mayan MH and Saheb-Chalan D. (1980). Observation on flock immunity to Newcastle disease after routine vaccination and experimental aerosol vaccination. Pol. Arch. Wete., (Warszawa). 21: 477-483.
Manchang TK, Abdu PA, Saidu L. (2004). Epidemiology and Clinicopathologic Manifestations of Newcastle Disease in Nigerian Local Chickens. Revue Élev. Méd. vét. Pays trop. 57: 35-39.
Manzoor R. (1999). Effective of Angovac (oil based killed HPS virus vaccine) against HPS in broilers. M.sc. Thesis, College of Veterinary Sciences, Lahore.
Nakamura K, Ohtsu N, Nakamura T, Yamamoto Y, Yamada M, Mase M, Imai K. (2008). Pathologic and immunohistochemical studies of Newcastle disease (ND) in broiler chickens vaccinated with ND: severe nonpurulent encephalitis and necrotizing pancreatitis. Vet Pathol. 45: 928-933.
http://dx.doi.org/10.1354/vp.45-6-928
PMid:18984798
Parry SH, Aitken ID. (1973). Immunoglobulin A in the respiratory tract of the chicken following exposure to Newcastle disease virus. Vet. Rec. 93: 258-260.
http://dx.doi.org/10.1136/vr.93.9.258
PMid:4760372
Salam R, Aslam A, Khan SA, Saeed K and Saleem G. (2003). Effect of different routes of vaccination against Newcastle disease on lymphoid organs of broilers. Pakistan. Vet. J.23 (2): 78-83
Susta L, Miller PJ, Afonso CL, Brown CC. (2011). Clinicopathological characterization in poultry of three strains of Newcastle disease virus isolated from recent outbreaks. Vet Pathol. 48: 349-360.
http://dx.doi.org/10.1177/0300985810375806
PMid:20685918
Tizard IR. (2004). Veterinary Immunology: An Introduction; 7th Ed.,W.B. Saunders Company, Philadelphia. 263-264.
Tizard IR. (1996). Veterinary Immunology. 5th Ed. W.B. Saunders Company, Philadelphia. 251-263.
Van-Eck JHH. (1987). Immunity to Newcastle Disease in fowl of different breeds primarily vaccinated with commercial inactivated oil-emulsion vaccines: a laboratory experiment. Vet. Q. 9: 296-303.
http://dx.doi.org/10.1080/01652176.1987.9694117
PMid:2962363
Voeten AC, Orthel FW, Jacobs J. (1977). Comparison of live Newcastle disease vaccines in a simple vaccination and challenge experiment. Res. Vet. Sci. 22: 138-145.
PMid:870956
Yuki Y and Kiyono H. (2003). New generation of mucosal adjuvants for the induction of protective immunity. Rev. Med. Virol. 13: 293–310.
http://dx.doi.org/10.1002/rmv.398
PMid:12931340
Zakeri A, Kashefi P. (2012). A Study of Programmed Cell Death in Cecal Tonsil of Infected Chickens by Infectious Bursal Disease Virus with Using Electronic Microscope. Am-Euras. J. Agric. & Environ. Sci. 12: 733-736.



