Journal of Infection and Molecular Biology
Review Article
Journal of Infection and Molecular Biology 1 (1): 1 – 8Proteins of Influenza Virus: a Review
Muhammad Abbas*, Zain ul Abidin
-
Veterinary Research Institute, Lahore 54600, Pakistan
*Corresponding author:qclabvri@yahoo.com
ARTICLE CITATION:
Abbas M, Abidin ZU (2013). Proteins of Influenza Virus: A Review. J. Inf. Mol. Biol. 1 (1): 1 – 8.
Received: 2013-02-27, Revised: 2013-03-09, Accepted: 2013-03-14
The electronic version of this article is the complete one and can be found online at
(
http://nexusacademicpublishers.com/table_contents_detail/2/17/html
)
which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited
ABSTRACT
Influenza virus infections are important both epidemically as well as pandemically and have great threat to avian species and humans especially, from last few decades. Certain pandemics of influenza virus have been reported causing severe damages. Once the host infected with influenza virus, it becomes immune-compromised and susceptible to secondary bacterial infections. Genome of influenza virus consists of eight segments of RNA, producing eleven types of proteins. Virulence and pathogenecity greatly depend upon its surface glycoproteins such as neuraminidase (NA) and hemagglutinin (HA). Other proteins of influenza virus are nucleoprotein (NP), 2 types of matrix proteins (M1 and M2), 2 non-structural proteins (NS1 and NS2), and polymerase subunit (PA) i.e. PB1, PB1-F2 and PB2. NP and M proteins are responsible for further classification of Influenza virus into influenza virus A, influenza virus B and influenza virus C. The aim of the review article is to discuss the structural importance of proteins of influenza A viruses.
INTRODUCTION
Influenza virus is involved in epidemic and pandemic diseases particularly respiratory system of both humans and animals. In 1918, influenza pandemically disturbed 50 million people and is still considered a great threat (Boivin et al., 2010). It belongs to family orthomyxoviridae and genus influenzavirus A. Influenza virus is an envelope, negative sense, single stranded virus with segmented RNA that produces 11 proteins (Hale et al., 2008). Waterfowls are considered to be the natural host for influenza A viruses (Jourdain et al., 2010). Virulence and pathogenecity of influenza virus greatly varies with its surface glycoproteins such as neuraminidase (NA) and hemagglutinin (HA) (Tumpey et al., 2005). NAs are nine in number while haemagglutinin proteins (HA) are sixteen in number (Fouchier et al 2005: Glaser et al., 2005: Yen et al., 2008). Other proteins of influenza A viruses include, nucleoprotein (NP), two types of matrix proteins i.e. M1 and M2, two non-structural proteins, i.e. NS1 and NS2 also called as nuclear export proteins (NEP), polymerase subunit (PA), PB1, PB1-F2 and PB2 (Ghedin et al., 2005). Based on the antigenic differences in NP and M, influenza viruses can be classified into three genera: influenza virus A, influenza virus B and influenza virus C. Two others, still not well accepted genera include isavirus and thogotavirus (Potter., 1998). Influenza A is mainly concerned with pandemic infections. It is reported that influenza viruses are pleomorphic in nature with mean diameter of 100–150 nm and the shape varies from filamentous to spherical (Roberts and Compans., 1998).
Certain pandemics have been reported most of which are; H1N1 Spanish flu in 1918, H2N2 Asian flu reported in Singapore in 1957, H2N2 epidemic in Japan in 1962 and Taiwan in 1964, H3N2 in Hong Kong flu in 1968 (Matrosovich et al., 2000; Shimada et al., 2009), H1N1 swine flu in Jersey in 1976, H1N1 Russian flu in USSR in 1977, H1N1 Swine flu in California in 2009 (Garten., et al., 2009: Smith et al.,2009). Influenza virus involved in immunosuppression was first time reported in 1908 (Kantzler et al., 1974). After infection with influenza virus, host becomes immune-compromised and susceptible to not only secondary bacterial infections but also other concurrent infections. Influenza virus damages the respiratory epithelial cells that ultimately result in invasion of secondary bacterial infections. Moreover, it was reported that Haemagllutinin (HA) was involved in inhibition of LPS-induced interleukin 12 (IL12) and p70 at transcriptional level making environment suitable for secondary bacterial infections (Noone et al., 2005). It has also been reported that influenza infections are fatal in patients having cardiac and pulmonary infection (Steininger et al., 2002). Influenza virus adapts different ways to circumvent host immune system, either by down regulation of host immune system or by enhancing its replication process to counteract with host immune system.
The basic purpose of this review article is to highlight the structural importance of proteins of influenza A viruses in relation to pathogenic standing, virulence and immunosuppression. Discussion will be made related to each protein of influenza A virus and their relationship with virulence; immune response of host, antigenic and pathogenic nature of virus.
LIFE CYCLE
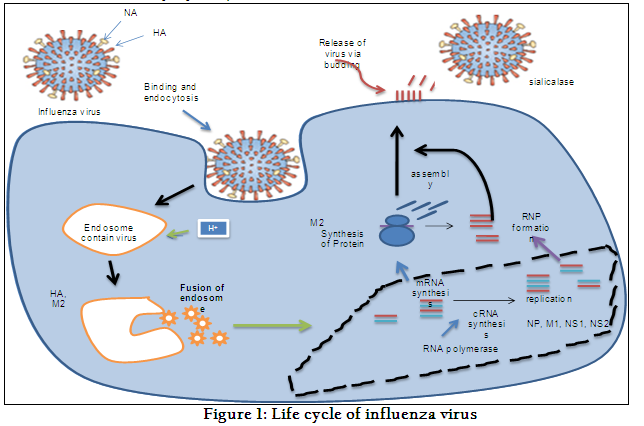
The detailed life cycle of influenza virus has been elaborated in Figure 1. There are three surface proteins of the influenza virus i.e. M2 protein, hemagglutinin (HA), and enzyme sialidase (NA). Influenza virus is an enveloped virus, it binds to α2-6 sialic acids (human influenza viruses) and to α 2-3 sialic acids (avian influenza viruses) via HA surface glycoproteins (Thompson et al., 2004). The enzymatic action of breaking α-ketosidic linkages by Neuraminidase (NA) (Hughes et al., 2000) leads the virus to enter into cell by endocytosis. This endocytosis occurs through receptor-mediated mechanism. After entry, virus binds with endosomal membrane. Endosome changes into lysosome due to decrease in pH up to 6 and HA molecule unfolds resulting into exposure of hidden hydrophobic chain of HA known as fusion peptide. Finally viral nucleocapsid releases into cytosole (Wharton et al., 1995) of the host cell. After release of nucleocapsid, protein synthesis occurs in cytoplasm. Finally virus releases via budding. sialidase elutes virus from cell surface and virus can now infect other cells (Itzstein, 2007).
HEMAGGLUTININ (HA)
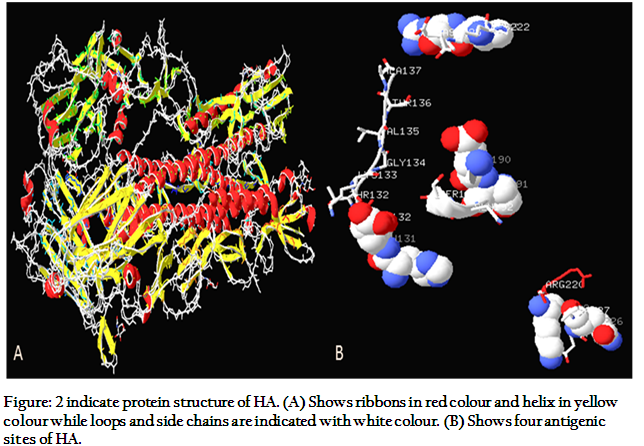
Influenza hemagglutinin (HA) is a glycoprotein, presented on the surface of virus. The molecular weight of HA is 165447.31 Dalton. Three chains H, J, L for polymer 1 with 328 amino acids and three chains I, K, M for polymer 2 with 160 amino acids have been reported. Extinction coefficient and theoretical pI have been calculated as 270060 and 6.15, respectively (Gamblin et al., 2004). HA protein has trimer structure, 135 Å at its tip. There are three subunits with two chains, so from final six chains, three are HA1s and three are HA2s. Monomer of HA1 consists of 328 residues, with elongated structure up to N-terminus of viral membrane; it ends after turning down at C-terminus. Distal portion of HA1 is jellyroll motif with eight anti-parallel beta sheets; α helix via forming loop separates jellyroll at position 3 and 4 forming pocket of sialic acid at top. Part of the distal end of HA1 forms an 8-stranded anti-parallel beta-sheet motif termed a "jelly roll." A short α-helix forms in the loop that separates 3rd and 4th strands of the jellyroll. HA2 consists of 221 residues with two anti-parallels α-helix having size of 75Å each. Single disulphide bond forms connection between HA1 and HA2. Whole molecule is stabilizing through van der Waals force (Wilson and Wiley., 1981).
There are four antigenic sites (Figure, 2) on hemagglutinin monomer; site A with 8 Å down from surface of molecule, site B binds externally to the residues of α-helix forming pocket or sialic acid binding, site C with 60 Å from lower tip of molecule, and site D with two β-sheets in jelly roll. Finally it can be concluded that any mutation in these antigenic sites can be changed in to virulent and antigenic properties of influenza virus (Schweiger et al., 2002). Influenza virus causes agglutination of red blood cells due to its hemagglutination ability. Based on which HA inhibition test is used for determination of virus titter (Madeley et al., 1971). The HA is also the receptor-binding site of the virus to initiate the infection as discussed before. Presently, 16 subtypes of HA are known ranging from H1 to H16. Recently, H16 was discovered from influenza A virus in black headed gulls in Sweden and Norway (Fouchier et al., 2005). The H1, H2, H3 are more prevalent in humans. H5N1 is more pathogenic in humans. It has also been studied that mutation in single amino acid of H5 of avian origin can change receptor acceptance to H5N1 in humans (Suzuki, 2005; Gambaryan et al., 2006).
NEURAMINIDASE (NA)
Three dimensional structure of NA at 2.2 Å indicates that it has single polypeptide chain A, four domains, with molecular weight of 46502.45 Dalton, Theoretical pI: 6.48 and Ext. coefficient of 85005 (Varghese and Colman, 1991). Structurally NA is a spikes square boxlike head and a stalk on surface of virus envelope. NA can also identify sialic acid (Itzstein et al., 1993) and has a role in catalyzation and cleavage of α-ketosidic linkage in terminal sialic acid residue and neighbouring residue of carbohydrate chain (Lentz and Air, 1986), so helping for the release of virus which results into spread of infection (Schulman and Palese., 1977). Active site of NA is present on its head (Colman et al., 1983). As a result of repeated infection, mutation occurs (Shu et al., 1993) due to which nine subtypes of NA has been developed (Kendal, 1987). There are two groups of NA: group 1 consists of, N1, N4, N5 and N8 while group 2 consists of N2, N3, N6, N7, and N9 (Russell et al., 2006).
The NA is glycoprotein and it is involved in antigenic variations such as antigenic drift or shift (Webster and Laver., 1975; Webster et al, 1982). The mutations have important role in circumventing immune responses. Such a shift was reported in NA of influenza A in 1957, which resulted into Asian influenza (H2N2) (Deshairs et al., 1986). Mutations in NA results into drug resistance so there is always need to improve drugs and vaccine with the passage of time (Thompson et al., 2004). Currently, there is an upsurge in the research to explore the possible mechanisms behind this resistance development especially for the drugs targeting the NA like Oseltamivir (TamiFlu).
NUCLEOPROTEINS (NP)
The NP is classified as viral nuclear proteins with molecular weight 170419.81 Dalton. It is a polypeptide nucleocapsid protein having one polymer, three chains i.e. A, B and C, with total length of 499 amino acids, theoretical pI: 9.24 and ext. coefficient of 167175. It is reported that at 3.2 Å, crystalline structure indicates that shape of protein is crescent like with a head, body and a flexible tail loop for attachment (Tarus et al., 2012). The RNA binding grove is present in the middle of head and body, outside the NP oligomer. There is a binding site for viral polymerase at body domain of NP. Furthermore tail loop can be a potent point for antiviral advancement (Ye et al., 2006). Eight RNA segments are rolled on NP subunits resulting formation of viral ribonucleoprotein (vRNP).
NP is positively charged, basic protein, present in 5th segment of mRNA (Hutchinson et al., 2009) and required for replication of virus (Huang et al., 1990). It is reported that using 32p orthophosphate labelling NP shows phosphorelation and has a role in viral replication and monitoring of viral yield (Kistner et al., 1989). Gene replacement shows that NP proteins are involved in host range determination (Altmuller et al., 1989) and site-specific phosphorylation (Kistner et al., 1985). It was studied on nucleoprotein primed mice that priming of NP could not stop infection but it can help in recovery via cross- reactive cytotoxic T cells (Wraith et al., 1987), CD8 T-cell gives cross-defence to influenza A (Zhong et al., 2010). Terminal part of 5th segment is involved in packaging of reporter genes. This transports between cytoplasm and nucleus via signals called nuclear localization signals (NLSs) (Neumann et al., 1997). Two NLSs have been reported for NP. NLS1 is conscientious for NP attachment to karyopherins α1 and α2 (Palese and Neill, 1997), found in residues 3 and 13. NLS1 is not only required for nuclear localization of NP but also involved in transport of vRNA to nucleus. NLS2 can be found at residues 198 to 216 in heart of NP and it functions as reporter protein and vRNA transcription (Weber et al., 1998). Moreover, NLS2 is essential for viral replication despite of NLS1 (Ozawa et al., 2007).
MATRIX (M) PROTEINS
M gene is responsible for matrix proteins including M1 and M2 proteins and membrane proteins. Segment 7 of influenza virus encodes for these M proteins. The size of M gene is 1027 bps. Furthermore M1 is found at 26 to 784 and M2 is located on nucleotide position 26 to 51 and 740 to 1007 (Lamb et al., 1981). These two proteins are discussed below.
M1 PROTEIN
The X-rays crystalline structure at pH 4.0, and 2.08-Å resolution of influenza A M1 protein indicates that N-terminal ranging amino acid from 2 to 158 is dimeric in shape with positive charge on its hydrophobic surface (Harris et al., 2001). The M1 protein is located at the inner side of virus in the shape of dimmer, creating a link among membrane proteins and inner core. The M1 has L domain motif that has important role in the assembly of viral part and budding of virus (Nayak et al., 2004). Gomez-Puertas et al., (2000) demonstrates that M1 proteins are involved in formation of virus-like particles i.e. VLPs instead of ribonucleoproteins. Function of M1 protein along with vRNA is not only the assembly of RNP in nucleus but it also transports it to cytoplasm (Huang et al., 2001). Latex agglutination test has been developed using chicken serum against M1 protein for identification of antibodies against influenza strains such as H3, H5, H7, and H9 (Wang et al., 2010).
M2 PROTEIN (The ion channel protein)
M2 protein has total 97 amino acids (Zebedee and Lamb, 1989) out of which there are five main amino acids at position 26, 27, 30, 31, and 34 that are important for drug resistance especially against adamantine (Liu et al., 2010). M2 is expressed on cell surface and integrated in virus. M2 is important for replication of virus at lower levels.
In nineties, it was believed that M2 protein is concerned in assembly of viral particles (Zebedee and Lamb, 1989), but latter it was also reported that M2 proteins was involved in regulation of ion channels particularly of monovalent ions. Ion channel function is concerned with acidification, which is vital phenomenon for virus entry. Hydrogen ions break linkage of vRNP and M1 resulting into RNP entry into cytoplasm after endocytosis of virus (Chen et al., 2008). Mutation in M2 protein can be resulted in drug resistance because of disturbance in ion channels regulation. Studies have shown that M2 protein forms trans-membrane domain. M2 ion channels are also affected by pH (Pinto et al., 1992). In Asia and USA anti-viral drug resistance against AIV has been reported due to single amino acid replacement i.e. serine to Asparagine (S31N) mutation in M2 protein (Nelson et al., 2009).
PB1, PB2 and PA PROTEINS (HA)
PB1 is located on segment 2 of the mRNA of influenza virus with two overlapping open reading frames (ORF) including PB1 and PB1-F2. These are located in mitochondria of eukaryotic cells and are responsible for intracellular localization of PB. It is also reported that another polypeptide is synthesized from second segment of mRNA named as PB1 N40 (Wise et al., 2009). PB1-F2 is made up of 90 amino acids, and is located in cytosol, nucleus and mitochondria especially in infected cells. It has been reported that PB1-F2 is involved in forming protein channels in mitochondrial membrane and for this function; c-terminal has great importance (Henkel et al., 2010). Khiabania et al., (2009) found that PB1 had a role in re-assortment events equally via inter and intra host. They also found that re-assortment patterns were same in pandemic influenza strains of 1957 and 1968 in humans. Human H2N2 and H3N2 re-assortment was only due to PB1 segment. Interaction of PB1 by its both N- and C-terminal with PA and PB2 resulting into heteromeric polymerase complex has been reported. Along with these traits, PB1 also has conserved RNA-dependent RNA polymerase motifs, viral RNA, complementary RNA, and nucleotide binding domain. Therefore, re-assortment and adaptation may be due to viral polymerase genes, which were later proved by Li et al., (2009). It was studied that PB1 may be absent in many influenza viruses leaving question mark on its importance in viral survival. So evolution of PB1 studies on truncated PB1 from different isolates of virus shows that it has effect on replication, and virulence but studies are needed to understand the effect of PB1-F2 on virus fitness (Trifonov et al., 2009). Structural weight of PB1-PA complex is 63530.73 Dalton. PB1-PA has two polymer chains, polymerase acidic protein polypeptide L chain A with 478 amino acids and RNA-directed RNA polymerase catalytic subunit polypeptide chain B with length of 81 amino acids (Obayashi et al., 2008).
The vRNA is catalyzed via complex processes with the help of viral RNA-dependent RNA polymerase that is made up of trimeric complex having three subunits, PB1, PB2 and PA in nucleus (Deng et al., 2005). PB2 has role in viral transcription via N terminal and is a cap binding protein (Gastaminza et al., 2003). Main protein is PB1 on which N-terminal is attached with PA and C-terminal is attached with PB2. One group of thought believed that PB1 and PA interacts in cytoplasm and then moves to nucleus with the help of RanBP5 while PB2 enters in nucleus, as monomer with help of Hsp90 (Momose et al., 2002) and then interacts with PB1 and PA complex, resulting in functional unit of PA-PB1-PB2 (Naito et al., 2007). Second opinion is that the PB1 and PB2 first intermingles in cytoplasm and enters in nucleus via Hsp90 protein and then PA protein attaches in the nucleus resulting in formation of functional complex but this concept is controversial. Hemerka et al., (2009) has developed a method called Bimolecular Fluorescence Complementation Assay (BiFC) which can analyze both PA-PB1 and PB1-PB2 interaction and have demonstrated that both are formed in cytoplasm and then travels to nucleus. They have also demonstrated that 100 amino acids on N-terminal region of PA protein are responsible for attachment to PB2 (Hemerka et al., 2009). PA, PB1, and PB2 are polymer in nature and along with NP; these are involved in forming ribonucleoprotines (RNP). RNPi further helps in RNA transcription, and replication (Ka-Leung et al., 2008)
Recent research shows that PB2 has role in inhibiting expression of interferon-β but its exact mechanism is unknown in addition of major virulence of influenza (Graef et al., 2010). PB2 is required for replication of virus and integral part of viral polymerase complex. PB2 has a role in determining host range and pathogenicity, proved by using squirrel monkeys, mice, mammalian cells, and pig (Manzoor et al., 2009). PA is phosphoprotein and functions as protease due to third amino terminal present closely to its nuclear localization signals. Mutations in PA result in down regulation of cRNA production from vRNA as polymerase action decreases.
NON-STRUCTURAL PROTEINS (STRUCTURE AND FUNCTIONS)
There are two non-structural proteins of influenza virus, i.e. NS1 and NS2 produced from 8th segment of RNA. These proteins are known as non-structural proteins as these proteins are produced in infected cell without integration into virion. These proteins are discussed in detailed as below.
NS1
NS1 protein has many functions with respect to virulence, importance in the disease pathogenesis, and studying host pathogen relationship. There are two domains of NS1 protein, i.e. N-terminal structural domain also known as RNA-binding domain (RBD) and effector domain (ED). The RBD has function in virus protection from host immune response via restricting cellular anti-viral proteins of special interest is 2'-5'-oligo (A) synthetase/RNase L pathway so inhibiting INFα/β production (Krug et al., 2003). Effector domain prevents development of host antiviral activity at cellular mRNAs level via preventing attachment and removal of mRNAs and polyadenylation specificity factor (CPSF) resulting in to inhibition of poly(A)-binding protein (PAB II) activity (Wang et al., 2002). It has also been reported that two domains of NS1 has a role in interaction with RNA and cellular proteins.
One most studied and important role of NS1 is to circumvent the host immune response via antagonizing IFN during viral replication. NS1 inhibits the production of IFN-inducible antiviral proteins: 2-5-oligoadenylate synthetase and dsRNA-dependent protein kinase R (PKR). dsRNA is required for the activation of 2´-5´OAS and PKR. The physical binding of NS1 to dsRNA results in inhibition of cellular 2-5 OAS and also in the inhibition of activation of PKR (Reviewed in Hale et al., 2006).
NUCLEAR AND CELLULAR LOCALIZATION OF NS1 PROTEIN
NS1 molecules travel between cytoplasm and nucleus of eukaryotic cells via nuclear pores. Nuclear localization depends on many factors including cell type, cell signals or nuclear localization signals (NLS), nuclear export signals (NES), time of exposure, and expression of NS1 protein, virus strain, and polarity of cells (Wen et al., 1995). Two nuclear localization signals have been reported for NS1 protein named as NLS1 and NLS2. NLS1 contains amino acids Asp-Arg-Leu-Arg-Arg codons from 34 to 38 and NLS2 is prevalent in region 203 and 237 amino acids (Greenspan et al., 1998).
NS1 has effect on nuclear post-transcriptional events either by inhibiting export of poly A mRNAs or via inhibiting splicing of pre mRNA. On the other hand, NS1 also effects viral mRNA translation in cytoplasm due to the binding of viral RNA with NS1 protein. This will affect phosphorylation of eukaryotic translation initiation factor 2α. NS1 protein contains NES which has similar hydrophobic spacing particularly leucine. Furthermore, replacement of alanine with leucine abrogates activity of nuclear export (Yongzhong et al., 1998).

NS2
NS2 is composed of 121 amino acids and translates from NS gene through mRNA splicing. It is reported that NS2 can be found in purified virus, and interacts with M1. NS2 along with M1 participates in viral RNPs nuclear export (Lommer and Luo, 2002; Iwatsuki-Horimoto et al., 2004), so NS2 is also known as nuclear export signal. Crm1 participates in this mechanism. Hence NS2 mediates interaction of M1 via its C-terminal and Crm1 by its N-terminal. In addition to these functions, it is also reported that NS2 alters RNA level resulting in decreased accumulation of transcription products and elevates the levels of viral replication hence it regulates viral transcription and translation (Robb et al., 2009).
EXPRESSION OF MX PROTEIN AND INTERFERON
Interferon alpha/beta (IFN-α/β) has relation with host defence system. There are three proteins involved in this system including protein kinase R, oligoadenylate synthetase, and Mx proteins (Stark et al., 1998). Mx protein is widely expressed as a response of viral infection in manyof the vertebrates. There are two types of Mx, including MxA and MxB found in nucleus and cytoplasm. It is reported that Mx protein and IFN-α/β shows its expression against influenza infection (Jung and Chae, 2006).
MECHANISM OF IMMUNITY IN INFLUENZA AND RECOVERY VIA CYTOKINES
In influenza virus infection, body response to combat the viral infection comes in the form of cell-mediated immunity. HA are main proteins involved in immunogenicity via humoral immunity. During influenza infection, antibodies against HA not only increase but also transfer to next generation passively, which ultimately enhance recovery. It is also reported that antibodies against NA have also a role in decreasing virus load and cell-to-cell proliferation. Experiments showed that antibodies produced against M1 and NP are not so important in recovery (Jakeman et al, 1989). Both HA and NA are responsible for neutralizing antibodies due to wide variations in their antigenic nature (Fu-Shi Quan et al., 2008).
It was believed that T cells are mainly involved in handling infections of influenza but experiments on mice proved that B cells were also involved in addition to T cells. So it is better to produce those vaccines that are capable to produce memory T cells and antibodies (Rangel-Moreno et al., 2010). Details of immune system activation and removal of influenza virus and infected cells have been described by Flint et al., (2004) and has been modified in figure 3.
CONCLUSIONS
Avian influenza occurrence is a reminder of our helplessness for emerging pandemic diseases. Vast verity of genetic shift and drift makes influenza virus dynamic, not only in causing infections but also producing mutations. Influenza virus contains eight segments of RNA, producing different eleven types of proteins. Every protein has unique characteristic and function in relation to virulence, pathogenic activity, and immunosuppression. Functional relationship of these proteins and immune system enables us to produce drugs and vaccines against mutated viruses like influenza virus. This review highlights the importance of influenza proteins in survival and development of virus with changing environment and its contribution in development of infection.
REFERENCES
Altmuller, Walter M, Fitch and Scholtissek C (1989). Biological and Genetic Evolution of the Nucleoprotein Gene of Human Influenza A Viruses. J. gen. Virol. 70: 2111-2119.
http://dx.doi.org/10.1099/0022-1317-70-8-2111
PMid:2769232
Boivin S, Cusack S, Ruigrok R and Hart D (2010). Influenza A Virus Polymerase: Structural Insights Into Replication And Host Adaptation Mechanisms. The American Society for Biochemistry and Molecular Biology. 10: 1-16.
Chen B, Leser G, Jackson D and Lamb R (2008). The Influenza Virus M2 Protein Cytoplasmic Tail Interacts with the M1 Protein and Influences Virus Assembly at the Site of Virus Budding. J. Virol. 82: 10059-10070.
http://dx.doi.org/10.1128/JVI.01184-08
PMid:18701586 PMCid:PMC2566248
Colman M, Varghese J and Laver W (1983). Structure of the catalytic and antigenic sites in influenza virus neuraminidase. Nature. 303: 41-44.
http://dx.doi.org/10.1038/303041a0
PMid:6188957
Deng T, Sharps J, Fodor E and Brownlee G (2005). In Vitro Assembly of PB2 with a PB1-PA Dimer Supports a New Model of Assembly of Influenza A Virus Polymerase Subunits into a Functional Trimeric Complex. J. Virol. 79: 8669–8674.
http://dx.doi.org/10.1128/JVI.79.13.8669-8674.2005
PMid:15956611 PMCid:PMC1143706
Deshairs C, Kessler N, Aymard M and Quash G (1986). Catalytic Properties of the A/H3N2 Influenza Neuraminidases: Influence of Antigenic Variations. J. gen. Virol. 67: 409 418.
http://dx.doi.org/10.1099/0022-1317-67-3-409
PMid:3950578
Flint J, Enquist W, Racaniello R and Skalka M (2004). Principles of virology. Molecular biology, pathogenesis, and control of animal viruses, ASM Press: Washington DC, USA, 2nd Edition.
Fouchier A, Munster V, Wallensten A, Bestebroer M, Herfst S, Smith D, Rimmelzwaan F, Olsen B and Osterhaus D (2005). Characterization of a novel influenza A virus hemagglutinin subtype (H16) obtained from black headed gulls. J. Virol.79: 2814–2822.
http://dx.doi.org/10.1128/JVI.79.5.2814-2822.2005
PMid:15709000 PMCid:PMC548452
Fu-Shi Quan, Compans R, Nguyen H and Kang SM (2008). Induction of Heterosubtypic Immunity to Influenza Virus by Intranasal Immunization. J. Virol. 82: 1350–1359.
http://dx.doi.org/10.1128/JVI.01615-07
PMid:18032492 PMCid:PMC2224423
Gambaryan A, Tuzikov A, Pazynina G, Bovin N, Balish A, Klimov A, (2006). Evolution of the receptor binding phenotype of influenza A (H5) viruses. J. Virol. 344: 432-438.
http://dx.doi.org/10.1016/j.virol.2005.08.035
PMid:16226289
Gamblin J, Haire F, Russell J, Stevens J, Xiao B, Ha Y, Vasisht N, Steinhauer A, Daniels S, Elliot A, Wiley C and Skehel J (2004). The structure and receptor binding properties of the 1918 influenza hemagglutinin. Science. 19: 1838-42.
http://dx.doi.org/10.1126/science.1093155
PMid:14764886
Gastaminza P, Perales B, Falco’n A and Ortín J (2003). Mutations in the N-Terminal Region of Influenza Virus PB2 Protein Affect Virus RNA Replication but Not Transcription. J. Virol. 77(9): 5098–5108.
http://dx.doi.org/10.1128/JVI.77.9.5098-5108.2003
PMid:12692212 PMCid:PMC153989
Ghedin E, Sengamalay N, Shumway M, Zaborsky J, Feldblyum T, Subbu V, Spiro D, Sitz J, Koo H, Bolotov P, Dernovoy D, Tatusova T, Bao Y, George K, Taylor J, Lipman D, Fraser C, Taubenberger J and Salzberg S (2005). Large-scale sequencing of human influenza reveals the dynamic nature of viral genome evolution. Nature. 437: 7062.
http://dx.doi.org/10.1038/nature04239
PMid:16208317
Glaser L, Stevens J, Zamarin D, Wilson I, García-Sastre A, Tumpey T, Basler C, Taubenberger J and Palese P (2005). A single amino acid substitution in 1918 influenza virus hemagglutinin changes receptor binding specificity. J. Virol. 79: 11533–11536.
http://dx.doi.org/10.1128/JVI.79.17.11533-11536.2005
PMid:16103207 PMCid:PMC1193621
Gómez-Puertas P, Albo C, Pérez-Pastrana E, Vivo A and Portela A (2000). Influenza Virus Matrix Protein Is the Major Driving Force in Virus Budding. J. Virol. 74: 11538–11547.
http://dx.doi.org/10.1128/JVI.74.24.11538-11547.2000
PMid:11090151 PMCid:PMC112434
Graef K, Vreede F, Lau Y, McCall A, Carr S, Subbarao K and Fodor E (2010). The PB2 subunit of the influenza virus RNA polymerase affects virulence by interacting with MAVS and inhibiting IFN- expression. J. Virol. 10: 879-90.
Greenspan D, Palese P and Krystal M (1998). Two Nuclear Location Signals in the Influenza Virus NS1 Nonstructural Protein. J. Virol. 62: 3020-3026.
Hale B, Jackson D, Chen Y, Lamb R and Randall R (2006). Influenza A virus NS1 protein binds p85β and activates phosphatidylinositol-3-kinase signaling. PNAS. 103: 14194–14199.
http://dx.doi.org/10.1073/pnas.0606109103
PMid:16963558 PMCid:PMC1599933
Hale B, Randall R Ortı’n J and Jackson D (2008). The multifunctional NS1 protein of influenza A viruses. J. gen. Virol. 89: 2359–2376.
http://dx.doi.org/10.1099/vir.0.2008/004606-0
PMid:18796704
Harris A, Forouhar F, Qiu S, Sha B and Luo M (2001). The crystal structure of the influenza matrix protein M1 at neutral pH: M1-M1 protein interfaces can rotate in the oligomeric structures of M1. International Congress Series, 1219: 405-410.
http://dx.doi.org/10.1016/S0531-5131(01)00349-1
Hemerka J, Wang D, Weng Y, Lu W, Kaushik R, Jin J, Aaron F, Harmon F and Li F (2009). Detection and Characterization of Influenza A Virus PA-PB2 Interaction through a Bimolecular Fluorescence Complementation Assay. J. Virol. 83: 3944–3955.
http://dx.doi.org/10.1128/JVI.02300-08
PMid:19193801 PMCid:PMC2663252
Henkel M, Mitzner D, Henklein P, Meyer-Almes F, Moroni A, DiFrancesco M, Henkes L, Kreim M, Kast S, Schubert U and Thiel G (2010). The Proapoptotic Influenza A Virus Protein PB1-F2 Forms a Nonselective Ion Channel. PLoS ONE. 5: e11112.
http://dx.doi.org/10.1371/journal.pone.0011112
PMid:20559552 PMCid:PMC2886074
Huang S, Palese P and Krystal M (1990) Determination of influenza virus proteins required for genome replication. J. Virol. 64: 5669–5673.
PMid:2214032 PMCid:PMC248627
Huang X, Liu T, Muller J, Levandowski A and Ye Z (2001). Effect of influenza virus matrix protein and viral RNA on ribonucleoprotein formation and nuclear export. Virology. 287: 405-16.
http://dx.doi.org/10.1006/viro.2001.1067
PMid:11531417
Hughes M, Matrosovich M, Rodgers E, Mcgregor M and Kawaoka Y (2000). Influenza A Viruses Lacking Sialidase Activity Can Undergo Multiple Cycles of Replication in Cell Culture, Eggs, or Mice. J. Virol. 74(11): 5206–5212.
http://dx.doi.org/10.1128/JVI.74.11.5206-5212.2000
PMid:10799596 PMCid:PMC110874
Hutchinson E, Wise H, Kudryavtseva K, Curran M and Digarda P (2009). Characterisation of influenza A viruses with mutations in segment 5 packaging signals. Vaccine. 27: 6270–6275.
http://dx.doi.org/10.1016/j.vaccine.2009.05.053
PMid:19840659 PMCid:PMC2771075
Itzstein M (2007). The war against influenza:discovery and development of sialidase inhibitors. Nature. 6: 967-973.
Itzstein M, Wu W, Kok g, Pegg M, Dyason J, Jin G, Phan T, Smythe M, White H, Oliver S, Colman P, Varghese J, Hotham, Cameron and Penn C (1993). Rational design of potent sialidase-based inhibitors of influenza virus replication. Nature. 363: 418 – 423.
http://dx.doi.org/10.1038/363418a0
PMid:8502295
Iwatsuki-Horimoto K, Horimoto T, Fujii Y and Kawaoka Y (2004). Generation of Influenza A Virus NS2 (NEP) Mutants with an Altered Nuclear Export Signal Sequence. J. Virol. 78: 10149–10155.
http://dx.doi.org/10.1128/JVI.78.18.10149-10155.2004
PMid:15331747 PMCid:PMC514982
Jakeman k, Smitht H and Sweet C (1989). Mechanism of Immunity to Influenza: Maternal and Passive Neonatal Protection Following Immunization of Adult Ferrets with a Live Vaccinia-Influenza Virus Haemagglutinin Recombinant but Not with Recombinants Containing Other Influenza Virus Proteins. J. Gen. Virol. 70: 1523-1531.
Jourdain E, Gunnarsson G, Wahlgren J, Latorre-Margalef N, Bro¨ jer C, Sahlin S, Svensson L, Waldenstro¨m J, Lundkvist A and Olsen B (2010). Influenza Virus in a Natural Host, the Mallard:Experimental Infection Data. Plos One. 5: e8935
http://dx.doi.org/10.1371/journal.pone.0008935
PMid:20126617 PMCid:PMC2812492
Jung K and Chae (2006). Expression of Mx Protein and Interferon-a in Pigs Experimentally Infected with Swine Influenza Virus. Vet Pathol. 43: 161–167.
http://dx.doi.org/10.1354/vp.43-2-161
PMid:16537933
Ka-Leung Ng A, Zhang H, Tan K, Li Z, Liu J, Chan P, Li S, Wing-Ngor Au S, Joachimiak A, Walz T, Wang J and Shaw P (2008). Structure of the influenza virus A H5N1 nucleoprotein: implications for RNA binding, oligomerization, and vaccine design. FASEB J. 22: 3638–3647.
http://dx.doi.org/10.1096/fj.08-112110
PMid:18614582 PMCid:PMC2537428
Kantzler G, lauteria S, Cusumano S, Lee J, Ganguly R and Waldman R (1974). Immunosuppression During Influenza Virus Infection. Infection and Immunity. 5: 996-1002.
Kendal A (1987). Epidemiologic implications of changes in the influenza virus genome. The American Journal of Medicine. 82: 4-14.
http://dx.doi.org/10.1016/0002-9343(87)90554-7
Khiabanian H, Trifonov V and Rabadan R (2009). Reassortment Patterns in Swine Influenza Viruses. PLoS Curr Influenza. 2: 1008.
Kistner O, Mlrller K and Scholtissek C (1985) Phosphopeptide Fingerprints of Nucleoproteins of Various Influenza A Virus Strains Grown in Different Host Cells. J. gen. Virol. 66: 465 472.
http://dx.doi.org/10.1099/0022-1317-66-3-465
PMid:3882883
Kistner O, Mlrller K and Scholtissek C (1989). Differential Phosphorylation of the Nucleoprotein of Influenza A Viruses. J. gen. Virol. 70: 2421-2431.
http://dx.doi.org/10.1099/0022-1317-70-9-2421
PMid:2778438
Krug M, Yuan W, Noah L and Latham G (2003). Intracellular warfare between human influenza viruses and human cells: The roles of the viral NS1 protein. Virology. 309: 181−189.
http://dx.doi.org/10.1016/S0042-6822(03)00119-3
Kumar N, Xin Z, Liang Y, Ly H and Liang Y (2008). NF-kB Signaling Differentially Regulates Influenza Virus RNA Synthesis. J. Virol. 82: 9880–9889.
http://dx.doi.org/10.1128/JVI.00909-08
PMid:18701591 PMCid:PMC2566266
Lamb R, Lait C and Choppin P (1981). Sequences of mRNAs derived from genome RNA segment 7 of influenza virus: Colinear and interrupted mRNAs code for overlapping proteins. Biochemistry. Proc. Natl Acad. Sci. USA. 78: 4170-4174.
http://dx.doi.org/10.1073/pnas.78.7.4170
PMid:6945577 PMCid:PMC319750
Lentz M and Air G (1986). Loss of enzyme activity in a site-directed mutant of influenza neuraminidase compared to expressed wild-type protein. Virology. 148: 74-83.
http://dx.doi.org/10.1016/0042-6822(86)90404-6
Li O, Chan M, Leung C, Chan R, Guan Y, Nicholls J and Poon L (2009). Full Factorial Analysis of Mammalian and Avian Influenza Polymerase Subunits Suggests a Role of an Efficient Polymerase for Virus Adaptation. PLoS ONE. 4: e5658.
http://dx.doi.org/10.1371/journal.pone.0005658
PMid:19462010 PMCid:PMC2680953
Liu H, Lv Y, Huang W, Yan M, Zhang W, Li M, Wang Q, Li J, Zheng D, Zhao Y, Sun C and Wang Z (2010). Detection of molecular markers of amantadine resistance in swine influenza viruses by pyrosequencing. Wei Sheng Wu Xue Bao. 50: 395-9.
PMid:20499646
Lommer B and Luo M (2002). Structural Plasticity in Influenza Virus Protein NS2 (NEP). The Journal of Biological Chemistry. 277: 7108–7117.
http://dx.doi.org/10.1074/jbc.M109045200
PMid:11751904
Madeley C, Allan W and Kendaly A (1971). Influenza A Viruses: Serological Relations of the Haemagglutinin and Neuraminidase Antigens of Ten Virus Isolates. J. gen. Virol. 12: 69-78.
http://dx.doi.org/10.1099/0022-1317-12-2-69
PMid:5166272
Manzoor R, Sakoda Y, Nomura N, Tsuda Y, Ozaki H, Okamatsu M and Kida H (2009). PB2 Protein of a Highly Pathogenic Avian Influenza Virus Strain A/chicken/Yamaguchi/7/2004 (H5N1) Determines Its Replication Potential in Pigs. J. Virol. 83: 1572–1578.
http://dx.doi.org/10.1128/JVI.01879-08
PMid:19052090 PMCid:PMC2643778
Matrosovich M, Tuzikov A, Bovin N, Gambaryan A, Klimov A, Castrucci R, Donatelli I and Kawaoka Y (2000). Early alterations of the receptor-binding properties of H1, H2, and H3 avian influenza virus hemagglutinins after their introduction into mammals. J. Virol. 74: 8502-12.
http://dx.doi.org/10.1128/JVI.74.18.8502-8512.2000
PMid:10954551 PMCid:PMC116362
Momose F, Naito T, Yano K, Sugimoto S, Morikawa Y and Nagata K (2002). Identification of Hsp90 as a Stimulatory Host Factor Involved in Influenza Virus RNA Synthesis. The Journal of Biological Chemistry. 277: 45306–45314.
http://dx.doi.org/10.1074/jbc.M206822200
PMid:12226087
Naito T, Momose F, Kawaguchi A and Nagata K (2007). Involvement of Hsp90 in assembly and nuclear import of influenza virus RNA polymerase subunits. J. Virol. 81: 1339–1349.
http://dx.doi.org/10.1128/JVI.01917-06
PMid:17121807 PMCid:PMC1797515
Nayak D, Ka-Wai Hui E and Barman S (2004). Assembly and budding of influenza virus. Virus Res. 106: 147–165.
http://dx.doi.org/10.1016/j.virusres.2004.08.012
PMid:15567494
Nelson M, Simonsen L, Viboud C, Miller M and Holmes E (2009). The origin and global emergence of adamantane resistant A/H3N2 influenza viruses. Virology. 388: 270–278.
http://dx.doi.org/10.1016/j.virol.2009.03.026
PMid:19394063 PMCid:PMC2705899
Neumann G, Castrucci M and Kawaoka Y (1997). Nuclear import and export of influenza virus nucleoprotein. J. Virol. 71: 9690–9700.
PMid:9371635 PMCid:PMC230279
Noone C, Lewis E, Frawely A, Newman R, hisMahon B, Mills K and Johnson P (2005). Novel mechanism of immunosuppression by influenza virus haemagglutinin: selective suppression of interleukin 12 p35 transcription in murine bone marrow-derived dendritic cells. J. gen. Virol. 86: 1885–1890.
http://dx.doi.org/10.1099/vir.0.80891-0
PMid:15958666
Obayashi E, Yoshida H, Kawai F, Shibayama N, Kawaguchi A, Nagata K, Tame R and Park Y (2008). The structural basis for an essential subunit interaction in influenza virus RNA polymerase. Nature. 454: 1127-31.
http://dx.doi.org/10.1038/nature07225
PMid:18660801
Ozawa M., Fujii K., Muramoto Y., Yamada S., Yamayoshi S., Takada A., Goto H., Horimoto T. and Kawaoka Y.(2007) Contributions of Two Nuclear Localization Signals of Influenza A Virus Nucleoprotein to Viral Replication. J. Virol. 81: 30–41.
http://dx.doi.org/10.1128/JVI.01434-06
PMCid:PMC1797272
Palese W, and Neill R. (1997). The NPI-1/NPI-3 (karyopherin α binding site on the influenza A virus nucleoprotein NP is a nonconventional nuclear localization signal. J. Virol. 71: 1850–1856.
PMid:9032315 PMCid:PMC191255
Pinto H, Holsinger J and Lamb A (1992) Influenza virus M2 protein has ion channel activity. Cell. 69: 517-28.
http://dx.doi.org/10.1016/0092-8674(92)90452-I
Potter W (1998). Chronicle of influenza pandemics In Textbook of Influenza. Black well Science. Oxford, UK.
Rangel-Moreno J, Carragher D, Misra R, Kusser K, Hartson L, Moquin A, Lund F and Randall T (2010). B Cells Promote Resistance to Heterosubtypic Strains of Influenza via Multiple Mechanisms. The Journal of Immunology. 180: 454-463.
http://dx.doi.org/10.4049/jimmunol.180.1.454
Rebecca J, Garten I, et al., (2009). Antigenic and Genetic Characteristics of Swine-Origin 2009 A (H1N1) Influenza Viruses Circulating in Humans. Science. 325: 197-201.
http://dx.doi.org/10.1126/science.1176225
PMid:19465683 PMCid:PMC3250984
Robb N, Smith M, Vreede F and Fodor E (2009). NS2/NEP protein regulates transcription and replication of the influenza virus RNA genome. J. gen. Virol. 90: 1398–1407.
http://dx.doi.org/10.1099/vir.0.009639-0
PMid:19264657
Roberts and Compans (1998) Host cell dependence of viral morphology. Microbiology. 95: 5746–5751.
Russell J, Haire F, Stevens J, Collins J, Lin P, Blackburn M, Hay J, Gamblin J and Skehel J (2006) The structure of H5N1 avian influenza neuraminidase suggests new opportunities for drug design. Nature. 443: 45-49.
http://dx.doi.org/10.1038/nature05114
PMid:16915235
Schulman J and Palese P (1977). Virulence factors of influenza A viruses: WSN virus neuraminidase required for plaque production in MDBK cells. J. Virol. 24: 170–176. PMid:561860 PMCid:PMC515920
Schweiger B, Zadow I and Heckler R (2002). Antigenic drift and variability of influenza viruses. Medical Microbiology and Immunology. 191: 133-138.
http://dx.doi.org/10.1007/s00430-002-0132-3
PMid:12458347
Shimada T, Kamiya H, Komiya N, Odaira F, Sunagawa T, Takahashi H, Toyokawa T, Tsuchihashi Y, Yasui Y, Tada Y and Okabe N (2009). Epidemiology Of Influenza A (H1N1) V Virus Infection In Japan, May - June 2009. Eurosurveillance. 14: 1-3.
Shu L, Bean W and Webster R (1993). Analysis of the evolution and variation of the human influenza A virus nucleoprotein gene from 1933 to 1990. J. Virol. 67: 2723–2729.
PMid:8474171 PMCid:PMC237595
Smith GJD, Vijaykrishna D, Bahl J, Lycett SJ, Worobey M, Pybus OG, Cheung CL, Raghwani J, Bhatt S, Peiris M, Guan Y and Rambaut A (2009). Origins and evolutionary genomics of the 2009 swine-origin H1N1 influenza A epidemicnear-final version. Nature, 459: 1122-1125.
http://dx.doi.org/10.1038/nature08182
PMid:19516283
Stark R, Kerr M, Williams G, Silverman H and Schreiber D (1998). how cells respond to interferons. Ann Rev Biochem. 67: 227–264.
http://dx.doi.org/10.1146/annurev.biochem.67.1.227
PMid:9759489
Steininger C, Holzmann H, Zwiauer F and Popow-Kraupp T (2002). Influenza A virus infection and cardiac arrhythmia during the neonatal period. Scand J Infect Dis.34: 782-784.
http://dx.doi.org/10.1080/00365540260348653
PMid:12477339
Suzuki Y (2005). Sialobiology of Influenza: Molecular Mechanism of Host Range Variation of Influenza Viruses. Biol. Pharm. Bull. 28: 399-408.
http://dx.doi.org/10.1248/bpb.28.399
PMid:15744059
Tarus B, Chevalier C, Richard C, Delmas B, Primo CD, Slama-Schwok A (2012). Molecular Dynamics Studies of the Nucleoprotein of Influenza A Virus: Role of the Protein Flexibility in RNA Binding. PLoS One 7(1): e30038.
http://dx.doi.org/10.1371/journal.pone.0030038
PMid:22272272 PMCid:PMC3260217
Thompson C, Barclay W and Zambon M (2004). Changes in in vitro susceptibility of influenza A H3N2 viruses to a neuraminidase inhibitor drug during evolution in the human host. J of Antimic Chemo. 53: 759–765.
http://dx.doi.org/10.1093/jac/dkh155
PMid:15028666
Trifonov V, Racaniello V and Rabadan R (2009). The Contribution of the PB1-F2 Protein to the Fitness of Influenza A Viruses and its Recent Evolution in the 2009 Influenza A (H1N1) Pandemic Virus. PLoS Curr Influenza. 3: RRN1006.
Tumpey T, Basler C, Aguilar P, Zeng H, Solórzano A, Swayne D, Cox N, Katz J, Taubenberger J, Palese P and García-Sastre A (2005). Characterization of the reconstructed 1918 Spanish influenza pandemic virus. Science. 310: 77-80.
http://dx.doi.org/10.1126/science.1119392
PMid:16210530
Varghese N and Colman M (1991). Three-dimensional structure of the neuraminidase of influenza virus A/Tokyo/3/67 at 2.2 A resolution. J Mol Biol. 221: 473-86.
http://dx.doi.org/10.1016/0022-2836(91)80068-6
Wang G, Hu S and Yu X (2010). Development of a latex agglutination test for detecting antibodies against avian influenza virus based on matrix 1 protein expressed in vitro. Avian Dis. 54: 41-5.
http://dx.doi.org/10.1637/9002-072309-Reg.1
PMid:20408397
Wang X, Basler F, Williams R, Silverman H, Palese P and Garcia-Sastre A (2002). Functional replacement of the carboxy-terminal two-thirds of the influenza A virus NS1 protein with short heterologous dimerization domains. J. Virol. 76: 12951−12962.
http://dx.doi.org/10.1128/JVI.76.24.12951-12962.2002
PMid:12438621 PMCid:PMC136679
Weber F, Kochs G, Gruber S and Haller O (1998). A classical bipartite nuclear localization signal on Thogoto and influenza A virus nucleoproteins. Virology. 250: 9–18.
http://dx.doi.org/10.1006/viro.1998.9329
PMid:9770415
Webster G, Laver G and SCHILD C (1982). Molecular mechanisms of variation in influenza viruses. Nature, 296: 115-121.
http://dx.doi.org/10.1038/296115a0
PMid:6174870
Webster O and Laver O (1975). Antigenic variation of influenza viruses. In The Influenza Viruses and Influenza. New York Accedimic Press, 269-314.
Weimin Zhong w, Liu F, Dong L, Lu X, Hancock K, Reinherz E, Katz J and Sambhara S (2010). Significant Impact of Sequence Variations in the Nucleoprotein on CD8 T Cell-Mediated Cross-Protection against Influenza A Virus Infections. PLoS One. 5: e10583.
http://dx.doi.org/10.1371/journal.pone.0010583
PMid:20485501 PMCid:PMC2868023
Wen W, Meinkoth L, Tsien Y and Taylor S (1995). Identification of a signal for rapid export of proteins from the nucleus. Cell. 82: 463-473.
http://dx.doi.org/10.1016/0092-8674(95)90435-2
Wharton S, Calder L, Ruigrokl R, Skehel J, Steinhauer D and Wiley D (1995). Electron microscopy of antibody complexes of influenza virus haemagglutinin in the fusion pH conformation. The EMBO Journal. 14: 240-246.
PMid:7835335 PMCid:PMC398077
Wilson A and Wiley S (1981). Structure of the haemagglutinin membrane glycoprotein of influenza virus at 3 Å resolution. Nature. 289: 366-373.
http://dx.doi.org/10.1038/289366a0
PMid:7464906
Wise H, Foeglein A, Sun J, Dalton R, Patel S, Howard W, Emma G, Barclay W and Digard P (2009). A Complicated Message: Identification of a Novel PB1-Related Protein Translated from Influenza A Virus Segment 2 mRNA. J. Virol. 83: 8021–8031.
http://dx.doi.org/10.1128/JVI.00826-09
PMid:19494001 PMCid:PMC2715786
Wraith D, Vessey A and Askonas B (1987). Purified Influenza Virus Nucleoprotein Protects Mice from Lethal Infection. J. gen. Virol. 68: 433-44.
http://dx.doi.org/10.1099/0022-1317-68-2-433
PMid:3493324
Ye Q, Krug M and Tao J (2006). The mechanism by which influenza A virus nucleoprotein forms oligomers and binds RNA. Nature. 444: 1078-82.
http://dx.doi.org/10.1038/nature05379
PMid:17151603
Yena H, Aldridgea J, Boona A, Ilyushinaa N, Salomona R, Hulse-Posta D, Marjukia H, Franksa J, Boltza D, Bushb D, Lipatova A, Webbya R, Rehgb J and Webstera R (2007). Changes in H5N1 influenza virus hemagglutinin receptor binding domain affect systemic spread. PNAS. 106: 286–291.
http://dx.doi.org/10.1073/pnas.0811052106
PMid:19116267 PMCid:PMC2629220
Yongzhong L, Yamakita Y and Krug R (1998). Regulation of a nuclear export signal by an adjacent inhibitory sequence: The effector domain of the influenza virus NS1 protein. Proc. Natl. Acad. Sci. USA. 95: 4864–4869.
http://dx.doi.org/10.1073/pnas.95.9.4864
Zebedee S and Lamb R (1989). Growth restriction of influenza A virus by M2 protein antibody is genetically linked to the M1 protein. Microbiol. 86: 1061-1065.



