Advances in Animal and Veterinary Sciences
Review Article
Advances in Animal and Veterinary Sciences 2 (4S): 17 – 23Special Issue–4 (2014): Reviews on Frontiers in Animal and Veterinary Sciences
West Nile Virus Infection among Animals and Humans in India
Baldev Raj Gulati1*, Akhil Kumar Gupta2, Surender Kumar Kadian2
- National Research Centre on Equines, Hisar, Haryana, India
- Department of Veterinary Microbiology, Lala Lajpat Rai University of Veterinary and Animal Sciences, Hisar, Haryana, India
*Corresponding author:brgulati@gmail.com
ARTICLE CITATION:
Gulati BR, Gupta AK, Kadian SK (2014). West Nile virus infection among animals and humans in India. Adv. Anim. Vet. Sci. 2 (4S): 17 – 23.
Received: 2014–04–15, Revised: 2014–04–22, Accepted: 2014–04–22
The electronic version of this article is the complete one and can be found online at
(
http://dx.doi.org/10.14737/journal.aavs/2014/2.4s.17.23
)
which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited
ABSTRACT
Since the introduction of West Nile Virus (WNV) in the New York City of USA in 1999, it has become a global public health concern. The outbreak of WNV has been reported in various parts of the world. The global spread of WNV has indicated that WNV is a re–emerging pathogen. Evidence of WNV infection in India is reported by sero–prevalence in 1952 and subsequently by virus isolation from mosquito, bat and man. Most of the WNV isolates from India belong to Lineage 5, while occasional isolates belonging to lineage 1a have also been reported, which are shown to be more virulent than those of lineage 5 isolates. WNV usually causes mild febrile illness but sometime neuro–invasive disorder has been reported. Currently, there is no licensed vaccine for human beings, although a number of veterinary vaccines have been licensed for horses. The information about prevalence of WNV among animal population in India is scanty. This review attempts to compile and analyze the information on WNV epidemiology, diagnosis and phylogeny from Indian perspective.
INTRODUCTION
West Nile Virus (WNV) is recognized as the most widespread zoonotic mosquito–transmitted arbovirus of public health importance (Pattan et al., 2009). It is a member of the genus Flavivirus and belongs to the Japanese encephalitis virus (JEV) antigenic complex under family Flaviviridae (De Filette et al., 2012). WNV was first isolated and identified in 1937 from the blood of a woman presented with mild febrile illness in the Nile district of Uganda (Smithburn et al., 1940). Since then many outbreaks of WNV have been reported in different parts of the world such as Egypt (1951), Israel (1957), France (1962–65), South Africa (1974), India (1980–81), Ukraine (1984), Algeria (1994), etc (Murgue et al., 2001). Until 1995, WNV was not considered an important human and animal pathogen as disease occurred only sporadically in humans and horses. However, first large outbreak of WNV was reported in Romania in 1996. The introduction of WNV in New York City of USA in 1999 has raised the major public health and veterinary concerns (Asnis et al., 2000; Roehrig, 2013). The New York 1999 (NY99) strain of WNV was first isolated from a dead American Crow (Lanciotti et al., 1999). It has been reported that 36% of the WNV infected horses either died of WNV or were euthanized. At present, WNV is widely prevalent in Africa, Southern Europe, Russia, Middle East, India, Australia and North America (Brandler and Tangy, 2013). From 1999 to 2010, more than 2.5 million people have been infected, with over 12,000 reported cases of encephalitis or meningitis and over 1,300 deaths (Kilpatrick, 2011). WNV infection is a self–limiting, non–fatal mild febrile illness causing mortality in horses, domestic and wild birds (Komar, 2000). This virus is known to be active in mosquitoes, birds and pigs (Paramasivan et al., 2003). WNV is reported to be maintained in the nature in a cycle involving birds and mosquitoes. Mosquitoes are principle vectors and birds act as both carriers and amplifying hosts for WNV. Animals and humans act as dead–end host (Paramasivan et al., 2003). In India, WNV infection has been reported from human (Balakrishnan et al., 2013; Chowdhury et al., 2014), animals (Ratho, et al., 1999) and birds (Mishra et al., 2012; Rodrigues et al., 1981). The purpose of this review is to review the status of WNV infection in India, both in human and animal population. The information on WNV epidemiology, diagnosis and phylogeny from Indian perspective has been reviewed in this article.
West Nile Virus Structure
WNV is a small spherical, enveloped particle containing single–stranded positive–sense non–segmented RNA genome having icosahedral symmetry. The genome is about 11 kb in size containing a single, long open reading frame flanked by non–coding regions at both ends (Brinton, 2002). Both the 5´ and 3´ non–coding regions of the genome form stem–loop structures that aid in replication, transcription, translation and packaging (Friebe and Harris, 2010). The viral RNA is translated as a single polyprotein that is post– and co–translationally cleaved by host and viral proteases, resulting in three structural (capsid, envelope and pre–membrane) and seven non–structural proteins (NS1, NS2A, NS2B, NS3, NS4A, NS4B and NS5) (Figure 1). The surface of virus particles contains two viral proteins; envelope protein (E, 53 kDa) and membrane protein (M, 8 kDa). The E glycoprotein is the major antigenic determinant on virus particles, mediates binding and fusion during virus entry. The M protein (8 kDa), produced during maturation of nascent virus particles within the secretory pathway, is a small proteolytic fragment of the precursor prM protein (18–20 kDa). Capsid protein (12 kDa) is involved in packaging of the viral genome and forming the nucleocapsid core (Mukhopadhyay et al., 2005).

Most neutralizing antibodies against flaviviruses recognize E protein. Crystallographic analysis of the soluble ectodomain of flavivirus E protein has revealed three domains (Modis et al., 2003; Rey et al., 1995). Domain I (DI) is the central domain and consists of an eight–stranded β – barrel that participates in the conformational changes associated with the acidification in the endosome. Domain II (DII) contains 12 β – strands and is formed from two extended loops that project from DI. DII is involved in the acid–catalyzed type II fusion event. The putative fusion peptide, which mediates insertion into the target cell membrane, is located at the tip of domain II distal to the transmembrane region. DIII adopts a seven–stranded immunoglobulin (Ig)–like fold and and exposes peptide loops with a role in cellular receptor binding and is a major target of neutralizing antibodies (Brinton, 2014).
Non–structural protein 1 (NS1) has both a “cellular” form and a “secreted” form; highly immunogenic and plays role in replication (Westaway et al., 2002). NS3 is a large (70 kDa) multifunctional protein having protease, nucleoside triphosphatase (NTPase) and helicase activity. This protein is responsible for cleaving other non–structural proteins from the viral polyprotein. The NS5 is a large (103 kDa) highly conserved multifunctional protein having RNA–dependent RNA polymerase (RdRP) and a methyltransferase (MTase) activity; it is necessary for viral replication (Lindenbach and Rice, 2003). Other non–structural proteins like NS2A, NS2B, NS4A and NS4B inhibit the component of innate immune response against virus infection and thus play a role in virus replication.
WNV Phylogenetic Diversity
Based on phylogenetic analysis, WNV has been previously classified into two major genetic lineages (Kramer et al., 2008). Lineage 1 is widespread in Europe, the USA, the Middle East, India, Africa and Australia while lineage 2 is mainly present in Sub–Sahran Africa, Madagascar and Central and Eastern Europe (Bakonyi et al., 2006; Platonov et al., 2008). In general, lineage 1 viruses are considered to be more virulent than the lineage 2 viruses. Lineage 1 is further subdivided into three different clades: 1a, 1b and 1c (De Filette et al., 2012). WNV–1a is mainly found in Europe, North America, Middle East and Africa (May et al., 2011). WNV–1b includes the Australian Kunjin virus and lineage 1c contains some Indian isolates.
Subsequently, based on sequence of C–prM–E region, five lineages of WNV have been established (Figure 2). Lineage 3 is represented by single virus strain isolated from mosquitoes in the Czech Republic, designated the Rabensburg virus (Bakonyi et al., 2005) and lineage 4 includes WNV isolated from Dermacentor marginatus ticks in the north–west Caucasus Mountain valley (Lvov et al., 2004). WNV strains from India belonging to a clade 1c of lineage 1 have now been classified as lineage 5 (Bondre et al., 2007). Three more putative lineages have been described. The Sarawak Kunjin virus strain is significantly different to the other Kunjin viruses and proposed as lineage 6, while the African virus (Koutango) has been proposed as lineage 7 (De Filette et al., 2012). WNV detected in Cx. pipiens mosquitoes captured in Spain have been proposed as lineage 8 (Vazquez et al., 2010).

WNV Epidemiology
The natural life cycle of WNV involves transmission from mosquitoes (primarily of the genus Culex) to wild birds (Figure 3). Birds act as amplifying host of the virus and the WNV has been isolated from nearly 300 species of birds. Mosquitoes become infected when they feed on infected birds that have high levels of viraemia (Pfeffer and Dobler, 2010). The virus then infects and replicate in the midgut of the mosquito (McGee et al., 2010; Richards et al., 2012). After replication in the midgut epithelium, the virus reaches the salivary glands from where it is transmitted to mammalian hosts during feeding (Colpitts et al., 2012; Suthar et al., 2013). Vertical transmission of WNV from female Culex pipiens mosquitoes to their progeny has been demonstrated in the laboratory (Bugbee and Forte, 2004). Transovarial transmission of WNV has also been experimentally demonstrated in Cx. vishnui mosquito (Mishra and Mourya, 2001). WNV can also be transmitted through blood transfusion, organ transplantation, breast–feeding and trans–placental transmission (
Sampatkumar, 2003).
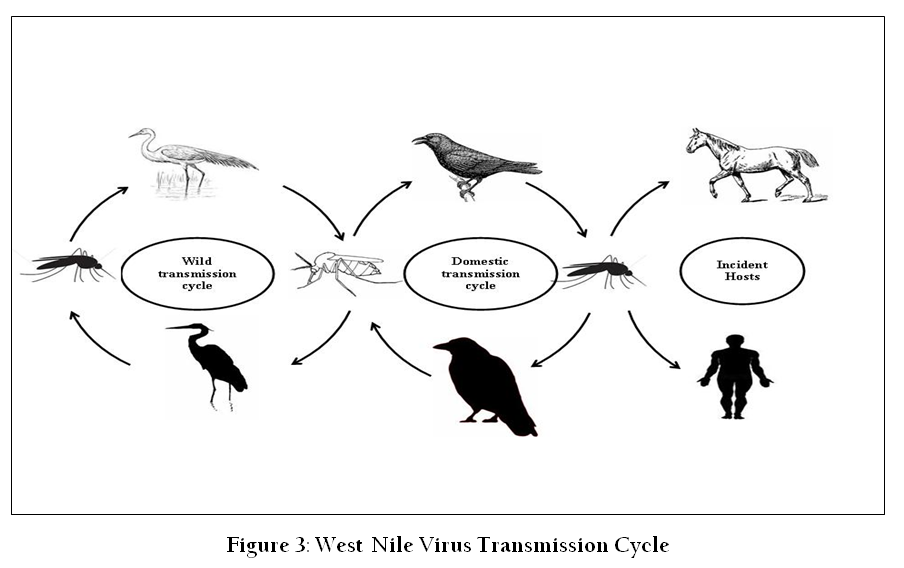
Ornithophilic mosquito species act as vectors for transmission of infection from viraemic birds to vertebrate hosts. The infected mosquito species vary according to geographical area. Culex mosquitoes are accepted as the primary global transmission vector (Hamer et al., 2008; Reisen et al., 2006; Vitek et al., 2008). There are reports of WNV transmission by Aedes mosquitoes, though they are not considered a primary vector in nature (Cupp et al., 2007; Farajollahi and Nelder, 2009; Vanlandingham et al., 2007). In India, Cx. tritaeniorhynchus, Cx. vishnui, Cx. bitaeniorhynchus and Cx. univittatus1, Cx. pipiens fatigans and Aedes albopictus have been reported as potential vectors of WNV (Ilkal et al., 1997; Varma, 1960).
WNV Prevalence in India
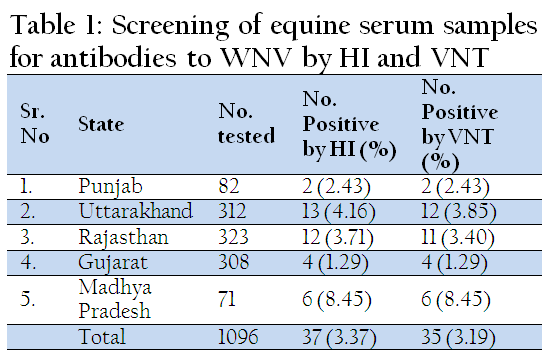
In India, seroprevalence of WNV in humans was reported from Mumbai by Banker (1952). WNV neutralizing antibodies have been detected in human sera collected from Tamil Nadu, Karnataka, Andhra Pradesh, Maharashtra, Gujarat, Madhya Pradesh, Orissa and Rajasthan (Paramasivan et al., 2003; Risbud et al., 1991; Rodrigues et al., 1980). During 1977, 1978 and 1981 serological evidence of WNV infections have been reported from Vellore and Kolar districts (George et al., 1984). Serological evidence against WNV has been reported among horses in and around Pune City (Parmasivan et al., 2003). In 2010, we screened 1096 equines from 5 states for WNV antibodies by haemagglutination inhibition (HI) and virus neutralization test (VNT). Of these 3.37% were tested positive for WNV antibodies by HI and 3.19% were found positive by VNT, which indicates circulation of WNV among Equine population (Table 1). In southern India, JEV/WNV neutralizing antibodies have been detected in ardeid birds mainly from the pond herons (Ardeola grayii) and cattle egrets (Bubulcus ibis), indicating their role in the natural cycle of WNV in India (Rodrigues et al., 1981).
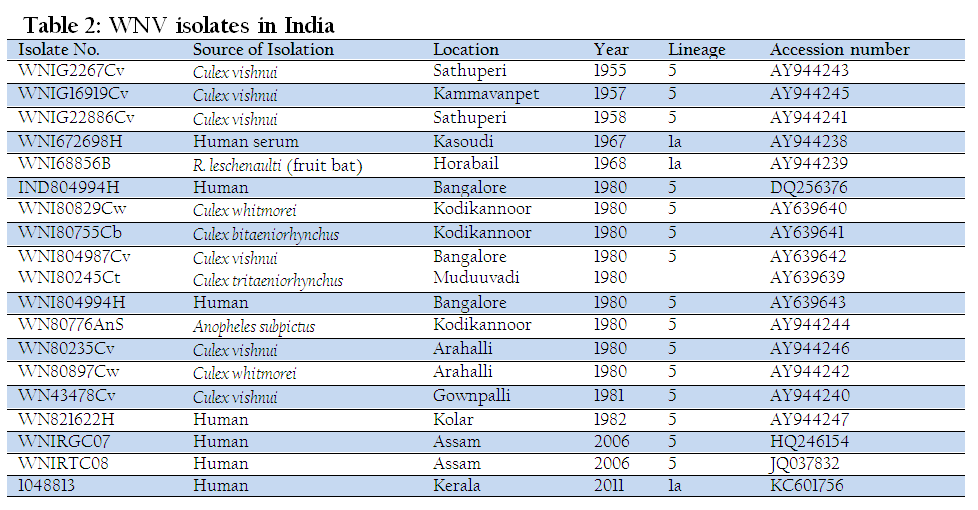
WNV strain P–4230 has been isolated from a laboratory worker who got infected while handling the Indian mosquito strain G–2266 and the Egyptian human strain E–101 (Paramasivan et al., 2003). In 2006, WNV belonging to lineage 5 has been detected among humans in the region of Assam to which JEV is endemic (Chowdhury et al., 2014; Khan et al., 2011). An outbreak of acute encephalitis syndrome (AES) due to WNV was reported in Kerala in 2011. The Kerala isolate has the highest nucleotide identity (96.2%) with the Russian isolate (GenBank Acc. no. DQ411030) and belong to Lineage 1 (clade 1a) (Balakrishnan et al., 2013). WNV has been isolated from human beings, frugivorous bat, domestic pigs and mosquitoes in India (Table 2) (George et al., 1984; Parmasivan et al., 2003; Paul et al., 1970; Rodrigues et al., 1980). However, WNV has not been isolated from horses in India.
WNV infection has been reported in pig population around Chandigarh city (Ratho, et al., 1999) and in wild migratory and resident water birds in Eastern and Northern India (Mishra et al., 2012). WNV has been isolated from various species of mosquito like Cx. tritaeniorhynchus, Cx. vishnui, Cx. Bitaeniorhynchus, etc (Table 1)
Diagnosis
Diagnosis of WNV infection is based largely on clinical signs, serological methods, nucleic acid based assays and virus isolation/antigen detection.
Serological Methods
Serological testing remains the primary method of diagnosing WNV infection, mainly based on anti–E antibody detection. However, cross–reactivity of the antibody response with related flaviviruses limits the specificity of serological tests. Plaque reduction neutralization test (PRNT) is the gold standard serological test for WNV detection (Dauphin and Zientara, 2007). It is used as a confirmatory method for detection of WNV–specific neutralizing antibodies from serum or cerebrospinal fluid (CSF). However, PRNT is a laborious test and must be carried out in a biosafety level 3 (BSL–3) facilities. HI test is used for the sero–prevalence of WNV antibodies in population. Demonstration of four–fold rise or drop of antibody titer in paired serum samples confirms the infection. But HI cannot differentiate between closely related flaviviruses and cannot be used as a confirmatory diagnostic method. Various enzyme–linked immunosorbent assays (ELISA) has been developed for WNV (Blitvich et al., 2003; Choi et al., 2007; Johnson et al., 2003; Kitai et al., 2007; Long et al., 2006). These tests have the advantage of being rapid, reproducible and less expensive. The IgM–antigen capture ELISA (MAC–ELISA) is optimal for the detection of early IgM antibodies, which allows an early diagnosis and the distinction between old and recent infections or vaccinations (De Filette et al., 2012). The MAC–ELISA enables the detection of acute infections since it detects early IgM antibodies in serum or CSF. The IgM antibodies can be detected within 4 to 7 days after the initial exposure and may persist for more than one year (Roehrig et al., 2003; Rossi et al., 2010). National institute of virology, Pune (India) developed MAC–ELISA kit for the diagnosis of WNV, Japanese Encephalitis (JE) and Dengue (DEN) viruses, which is commercialized by Zydus Cadila. We developed a panel of murine monoclonal antibodies (MAbs) against WNV and using these mAbs, developed a blocking–ELISA for epitope detection of WNV in Horses (Gupta et al., 2013).
Nucleic Acid Based Assays
WNV can be detected by conventional reverse–transcription polymerase chain reaction (RT–PCR), real–time PCR and multiplex reverse transcription–PCR ligase detection reaction (Johnson et al., 2001; Linke et al., 2007; Rondini et al., 2008; Shi and Kramer, 2003). A SYBR Green based assay that could detect 100% of the different WNV target region variants have been developed (Papin et al., 2004). But this SYBR based assay has lower sensitivity as compared to WNV–specific TaqMan RT–PCR assays (Johnson et al., 2010). The two most popular alternatives to SYBR Green are TaqMan and molecular beacons, both of which use hybridization probes and rely on fluorescence resonance energy transfer (FRET) for quantification (Jimenez–Clavero et al., 2006; Lanciotti et al., 2000). Parida and co–workers (2004) developed a loop–mediated isothermal amplification (LAMP) assay for WNV, which is relatively inexpensive. In India during 2010–11, conventional RT–PCR, real–time RT–PCR and RT–LAMP assays were used to detect WNV associated with multifocal retinitis in patients from southern India (Shukla et al., 2012).
Antigen Detection/Virus Isolation
It is the method of choice for detection of WNV in vertebrate/ mosquito pools/avian samples. Virus can be isolated from CSF, blood or brain tissues in cell cultures viz: Vero/BHK 21/RK–13/mosquito (AP61) cell lines (Castillo–Olivares and Wood, 2004). Antigen capture ELISA has been developed for the detection of WNV using NS–1 (Chung and Diamond, 2008; Macdonald et al., 2005). A membrane–based electrochemical nano–biosensor has been developed for the detection of WNV that recognizes viral particles or virus E protein during the early stage of infection (Nguyen et al., 2009). In India, WNV has been isolated by intra–cerebral inoculation in 3–day old infant mice; inoculation in BHK–21 and Vero cell lines (Balakrishnan et al., 2013; Bondre et al., 2007; Chowdhury et al., 2014).
Vaccination
Vaccination is the primary method of reducing the risk of infection from WNV. At present, there are no commercially available vaccines for human use. A number of vaccine strategies have been explored and some human candidate vaccines have been evaluated in Phase I and II clinical trials (Table 3) (Iyer and Kousoulas, 2013).
Compared to WNV vaccine for human, there are effective licensed vaccines for horses (De Filette et al., 2012). Fort Dodge Animal Health (subsidiary of Pfizer) developed a vaccine under the trade name West Nile–Innovator® which is formalin–inactivated whole West Nile virus (Ng et al., 2003). This vaccine is currently commercialized in the USA and is quite effective. Another killed virus vaccine (Vetera® WNV vaccine) developed by Boehringer Ingelheim Vetmedica is also licensed by the United States Department of Agriculture (USDA). A third commercialized WNV vaccine in the United States for horses is Recombitek® Equine West Nile Virus Vaccine (Merial, now Sanofi Aventis), which is a chimeric recombinant canarypoxvirus vaccine (El Garch et al., 2008). In 2005, Fort Dodge Animal Health/Pfizer developed a DNA based WNV vaccine under the trade name of West Nile–Innovator® DNA. The vaccine contains an unformulated plasmid DNA encoding the prM and E protein of WNV and MetaStim™ as adjuvant. This vaccine has recently been discontinued by Pfizer (Brandler and Tangy, 2013).
CONCLUDING REMARKS
WNV infection is emerging as an important infection of human and animals across the globe. In India, WNV infection has been reported from humans, pigs, bats and birds. A few studies indicate the seroprevalence of WNV among horses in India. Mosquitoes of mainly Culex species and sometime Anopheles subpictus are the main vectors of WNV in India. Some data has emerged during last 5 decades on epidemiology of WNV in India. However, there is a need for systematic study to know the status of WNV infection in human and different animals, particularly horses in India. Role of mosquitoes and various birds in spread of WNV in newer geographical regions of India also needs further investigations.
Research has gained pace regarding the public health significance of the WNV infection. However, there is greater amount of serological cross–reactivity of WNV with related flavivirues circulating in a region. For instance, in India WNV and Japanese encephalitis virus (JEV) both are endemic and co–circulate in human and animal population. Therefore, there is need for development of sensitive and rapid sero–diagnostics, which can clearly discriminate WNV from JEV in India. Development of sensitive and rapid molecular diagnostics should be encouraged for early detection of infection. The environmental factors play critical role in emergence, maintenance and transmission of the WNV to newer geographical regions. Hence, impact of changing environment in the ecology of WNV needs to be investigated. There is no specific treatment for WNV; therefore strong surveillance system needs to be developed for understanding the pattern of disease distribution. Efforts for development of immuno–prophylactics for human and animals should be done for control of WNV infection. It is hoped that the increase in knowledge of interaction of virus, vector and host will provide new insights for the therapeutic and preventive measures.
ACKNOWLEDGMENTS
The authors thank Indian Council of Agricultural Research and National Research Centre on Equines for the support for the study.
CONFLICT OF INTEREST
The authors declare no conflict of interest.
REFERENCES
Asnis DS, Conetta R, Teixeira AA, Waldman G, Sampson BA (2000). The West Nile virus outbreak of 1999 in New York: the Flushing Hospital experience. Clin. Infect. Dis. 30: 413–418.
http://dx.doi.org/10.1086/313737
PMid:10722421
Bakonyi T, Hubalek Z, Rudolf I, Nowotny N (2005). Novel flavivirus or new lineage of West Nile virus, Central Europe. Emerg. Infect. Dis. 11: 225–231.
http://dx.doi.org/10.3201/eid1102.041028
PMid:15752439 PMCid:PMC3320449
Bakonyi T, Ivanics E, Erdelyi K, Ursu K, Ferenczi E, Weissenbock H, Nowotny N (2006). Lineage 1 and 2 strains of encephalitic West Nile virus, Central Europe. Emerg. Infect. Dis. 12: 618–623.
http://dx.doi.org/10.3201/eid1204.051379
PMid:16704810 PMCid:PMC3294705
Balakrishnan A, Butte DK, Jadhav SM (2013). Complete genome sequence of west nile virus isolated from alappuzha district, kerala, India. Genome Announc. 16: 1.
Banker DD (1952). Preliminary observations on antibody patterns against certain viruses among inhabitants of Bombay city. Indian J. Med. Sci. 6: 733–746.
Blitvich BJ, Bowen RA, Marlenee NL, Hall RA, Bunning ML, Beaty BJ (2003). Epitope–Blocking Enzyme–Linked Immunosorbent Assays for detection of West Nile Virus antibodies in domestic mammals. J. Clin. Microbiol. 41: 2676–2679.
http://dx.doi.org/10.1128/JCM.41.6.2676-2679.2003
http://dx.doi.org/10.1128/JCM.41.3.1041-1047.2003
PMid:12791902
Bondre VP, Jadi RS, Mishra AC, Yergolkar PN, Arankalle VA (2007). West Nile virus isolates from India: evidence for a distinct genetic lineage. J. Gen. Virol. 88: 875–884.
http://dx.doi.org/10.1099/vir.0.82403-0
PMid:17325360
Brandler S, Tangy F (2013). Vaccines in Development against West Nile Virus. Viruses 5: 2384–2409.
http://dx.doi.org/10.3390/v5102384
PMid:24084235 PMCid:PMC3814594
Brinton MA (2002). The molecular biology of West Nile Virus: a new invader of the western hemisphere. Annu. Rev. Microbiol. 56: 371–402.
http://dx.doi.org/10.1146/annurev.micro.56.012302.160654
PMid:12142476
Brinton MA (2014). Replication Cycle and Molecular Biology of the West Nile Virus. Viruses 6: 13–53.
http://dx.doi.org/10.3390/v6010013
PMid:24378320 PMCid:PMC3917430
Bugbee LM, Forte LR (2004). "The discovery of West Nile virus in overwintering Culex pipiens (Diptera: Culicidae) mosquitoes in Lehigh County, Pennsylvania". J. Am. Mosq. Control Assoc. 20: 326–327.
PMid:15532939
Castillo–Olivares J, Wood J (2004). West Nile virus infection of horses. Vet. Res. 35: 467–483.
http://dx.doi.org/10.1051/vetres:2004022
PMid:15236677
Choi K, Ko Y, Nah J, Kim Y, Kang S,Yoon K, Joo Y (2007). Monoclonal Antibody–Based Competitive Enzyme–Linked Immunosorbent Assay for Detecting and Quantifying West Nile Virus–Neutralizing Antibodies in Horse Sera. Clin. Vaccine Immunol. 14: 134–138.
http://dx.doi.org/10.1128/CVI.00322-06
PMid:17135450 PMCid:PMC1797797
Chowdhury P, Khan SA, Dutta P, Topno R, Mahanta J (2014). Characterization of West Nile virus (WNV) isolates from Assam, India: insights into the circulating WNV in northeastern India. Comp. Immunol. Microbiol. Infect. Dis. 37: 39–47.
http://dx.doi.org/10.1016/j.cimid.2013.10.006
PMid:24268432
Chung KM, Diamond MS (2008). Defining the levels of secreted non–structural protein NS1 after West Nile virus infection in cell culture and mice. J. Med. Virol. 80: 547–556.
http://dx.doi.org/10.1002/jmv.21091
PMid:18205232 PMCid:PMC2696118
Colpitts TM, Conway MJ, Montgomery RR, Fikrig E (2012). West nile virus: biology, transmission, and human infection. Clin. Microbiol. Rev. 25: 635–648.
http://dx.doi.org/10.1128/CMR.00045-12
PMid:23034323 PMCid:PMC3485754
Cupp EW, Hassan HK, Yue X, Oldland WK, Lilley BM, Unnasch TR (2007). West Nile virus infection in mosquitoes in the mid–south USA, 2002–2005. J. Med. Entomol. 44: 117–125.
http://dx.doi.org/10.1603/0022-2585(2007)44[117:WNVIIM]2.0.CO;2
Dauphin G, Zientara S (2007). West Nile virus: Recent trends in diagnosis and vaccine development. Vaccine 25: 5563–5576.
http://dx.doi.org/10.1016/j.vaccine.2006.12.005
PMid:17292514
De Filette M, Ulbert S, Diamond M, Sanders NN (2012). Recent progress in West Nile virus diagnosis and vaccination. Vet. Res. 43: 16.
http://dx.doi.org/10.1186/1297-9716-43-16
PMid:22380523 PMCid:PMC3311072
El Garch H, Minke JM, Rehder J, Richard S, Edlund Toulemonde C, Dinic S, Andreoni C, Audonnet JC, Nordgren R, Juillard V (2008). A West Nile virus (WNV) recombinant canarypox virus vaccine elicits WNV–specific neutralizing antibodies and cell–mediated immune responses in the horse. Vet. Immunol. Immunopathol. 123: 230–239.
http://dx.doi.org/10.1016/j.vetimm.2008.02.002
PMid:18372050
Farajollahi A, Nelder MP (2009). Changes in Aedes albopictus (Diptera: Culicidae) populations in New Jersey and implications for arbovirus transmission. J. Med. Entomol. 46: 1220 –1224.
http://dx.doi.org/10.1603/033.046.0533
PMid:19769057
Friebe P, Harris E (2010). Interplay of RNA elements in the dengue virus 5´ and 3´ ends required for viral RNA replication. J. Virol. 84: 6103– 6118.
http://dx.doi.org/10.1128/JVI.02042-09
PMid:20357095 PMCid:PMC2876622
George S, Gourie–Devi M, Rao JA, Prasad SR, Pavri KM (1984). Isolation of West Nile virus from the brains of children who had died of encephalitis. Bull WHO. 62: 879–882.
PMid:6099760 PMCid:PMC2536262
Gupta AK, Kadian SK, Gulati BR (2013). Production and Characterization of Monoclonal Antibodies against West Nile Virus. Vet. Pract. 14: 275–278.
Hamer GL, Kitron UD, Brawn JD, Loss SR, Ruiz MO, Goldberg TL, Walker ED (2008). Culex pipiens (Diptera: Culicidae): a bridge vector of West Nile virus to humans. J. Med. Entomol. 45: 125–128.
http://dx.doi.org/10.1603/0022-2585(2008)45[125:CPDCAB]2.0.CO;2
Ilkal MA, Mavale MS, Prasanna Y, Jacob PG, Geevarghese G, Banerjee K (1997). Experimental studies on the vector potential of certain Culex species to West Nile virus. Indian J. Med. Res. 106: 225–228.
PMid:9378528
Iyer AV, Kousoulas KG (2013). A Review of Vaccine Approaches for West Nile Virus. Int. J. Environ. Res. Public Health 10: 4200–4223.
http://dx.doi.org/10.3390/ijerph10094200
PMid:24025396 PMCid:PMC3799512
Jimenez–Clavero MA, Aguero M, Rojo G, Gomez–Tejedor C (2006). A new fluorogenic real–time RT–PCR assay for detection of lineage 1 and lineage 2 West Nile viruses. J. Vet. Diagn. Invest. 18: 459–462.
http://dx.doi.org/10.1177/104063870601800505
PMid:17037613
Johnson AJ, Langevin S, Wolff KL, Komar N (2003). Detection of Anti–West Nile Virus Immunoglobulin M in Chicken Serum by an Enzyme–Linked Immunosorbent Assay. J. Clin. Microbiol. 41: 2002–2007.
http://dx.doi.org/10.1128/JCM.41.5.2002-2007.2003
PMid:12734241 PMCid:PMC154686
Johnson DJ, Ostlund EN, Pedersen DD, Schmitt BJ (2001). Detection of North American West Nile virus in animal tissue by a reverse transcription–nested polymerase chain reaction assay. Emerg. Infect. Dis. 7: 739–741.
http://dx.doi.org/10.3201/eid0704.017425
http://dx.doi.org/10.3201/eid0704.010425
PMid:11585541 PMCid:PMC2631755
Johnson N, Wakeley PR, Mansfield KL, McCracken F, Haxton B, Phipps LP, Fooks AR (2010). Assessment of a novel real–time pan–flavivirus RT–polymerase chain reaction. Vector Borne Zoonotic Dis. 10: 665–671.
http://dx.doi.org/10.1089/vbz.2009.0210
PMid:20854019
Khan SA, Dutta P, Khan AM, Chowdhury P, Borah J, Doloi P, Mahanta J (2011). West Nile virus infection, Assam, India. Emerg. Infect. Dis. 17: 947–948.
http://dx.doi.org/10.3201/eid1705.100479
PMid:21529426 PMCid:PMC3321754
Kilpatrick AM (2011). Globalization, land use, and the invasion of West Nile virus. Science 334: 323–327.
http://dx.doi.org/10.1126/science.1201010
PMid:22021850 PMCid:PMC3346291
Kitai Y, Shoda M, Kondo T, Konishi E (2007). Epitope–Blocking Enzyme–Linked Immunosorbent assay to differentiate West Nile Virus from Japanese Encephalitis Virus Infections in Equine Sera. Clin. Vaccine Immunol. 14: 1024–1031.
http://dx.doi.org/10.1128/CVI.00051-07
PMid:17596430 PMCid:PMC2044481
Komar N (2000). West Nile viral encephalitis. Rev. Sci. Tech. 19: 166–176.
PMid:11189714
Kramer LD, Styer LM, Ebel GD (2008). A global perspective on the epidemiology of West Nile virus. Annu. Rev. Entomol. 53: 61–81.
http://dx.doi.org/10.1146/annurev.ento.53.103106.093258
PMid:17645411
Lanciotti RS, Roehrig JT, Deubel V, Smith J, Parker M, Steele K, Crise B, Volpe KE, Crabtree MB, Scherret JH, Hall RA, MacKenzie JS, Cropp CB, Panigrahy B, Ostlund E, Schmitt B, Malkinson M, Banet C, Weissman J, Komar N, Savage HM, Stone W, McNamara T, Gubler DJ (1999). Origin of the West Nile virus responsible for an outbreak of encephalitis in the northeastern United States. Science 286: 2333-2337.
http://dx.doi.org/10.1126/science.286.5448.2333
PMid:10600742
Lanciotti RS, Kerst AJ, Nasci RS, Godsey MS, Mitchell CJ, Savage HM, Komar N, Panella NA, Allen BC, Volpe KE, Davis BS, Roehrig JT (2000). Rapid detection of west nile virus from human clinical specimens, field collected mosquitoes, and avian samples by a TaqMan reverse transcriptase–PCR assay. J. Clin. Microbiol. 38: 4066–4071.
PMid:11060069 PMCid:PMC87542
Lindenbach BD, Rice CM (2003). Molecular biology of flaviviruses. Adv. Virus Res. 59: 23– 61.
http://dx.doi.org/10.1016/S0065-3527(03)59002-9
Linke S, Ellerbrok H, Niedrig M, Nitsche A, Pauli G (2007). Detection of West Nile virus lineages 1 and 2 by real–time PCR. J. Virol. Methods. 146: 355–358.
http://dx.doi.org/10.1016/j.jviromet.2007.05.021
PMid:17604132
Long MT, Jeter W, Hernandez J, Sellon DC, Gosche D, Gillis K, Bille E, Gibbs EP (2006). Diagnostic performance of the equine IgM capture ELISA for serodiagnosis of West Nile virus infection. J. Vet. Intern. Med. 20: 608–613.
http://dx.doi.org/10.1111/j.1939-1676.2006.tb02904.x
PMid:16734097
Lvov DK, Butenko AM, Gromashevsky VL, Kovtunov AI, Prilipov AG, Kinney R, Aristova VA, Dzharkenov AF, Samokhvalov EI, Savage HM, Shchelkanov MY, Galkina IV, Deryabin PG, Gubler DJ, Kulikova LN, Alkhovsky SK, Moskvina TM, Zlobina LV, Sadykova GK, Shatalov AG, Lvov DN, Usachev VE, Voronina AG (2004). West Nile virus and other zoonotic viruses in Russia: examples of emerging–reemerging situations. Arch. Virol. Suppl. 18: 85–96.
PMid:15119764
Macdonald J, Tonry J, Hall RA, Williams B, Palacios G, Ashok MS, Jabado O, Clark D, Tesh RB, Briese T, Lipkin WI (2005). NS1 protein secretion during the acute phase of West Nile virus infection. J. Virol. 79: 13924–13933.
http://dx.doi.org/10.1128/JVI.79.22.13924-13933.2005
PMid:16254328 PMCid:PMC1280181
May FJ, Davis CT, Tesh RB, Barrett AD (2011). Phylogeography of West Nile virus: from the cradle of evolution in Africa to Eurasia, Australia, and the Americas. J. Virol. 85: 2964–2974.
http://dx.doi.org/10.1128/JVI.01963-10
PMid:21159871 PMCid:PMC3067944
McGee CE, Shustov AV, Tsetsarkin K, Frolov IV, Mason PW, Vanlandingham DL, Higgs S (2010). Infection, dissemination, and transmission of a West Nile virus green fluorescent protein infectious clone by Culex pipiens quinquefasciatus mosquitoes. Vector Borne Zoonotic Dis. 10: 267–274.
http://dx.doi.org/10.1089/vbz.2009.0067
PMid:19619041 PMCid:PMC2883532
Mishra AC, Mourya DT (2001). Transovarial transmission of West Nile virus in Culex vishnui mosquito. Ind. J. Med. Res. 114: 212–214.
PMid:12040765
Mishra N, Kalaiyarasu S, Nagarajan S, Rao MV, George A, Sridevi R, Behera SP, Dubey SC, McCracken T, Newman SH (2012). Serological evidence of West Nile virus infection in wild migratory and resident water birds in Eastern and Northern India. Comp. Immunol. Microbiol. Infect. Dis. 35: 591–598.
http://dx.doi.org/10.1016/j.cimid.2012.08.002
PMid:22925932
Modis Y, Ogata S, Clements D, Harrison SC (2003). A ligand–binding pocket in the dengue virus envelope glycoprotein. Proc. Natl. Acad. Sci. USA 100: 6986–6991.
http://dx.doi.org/10.1073/pnas.0832193100
PMid:12759475 PMCid:PMC165817
Mukhopadhyay S, Kuhn RJ, Rossmann MG (2005). A Structural Perspective of the Flavivirus life cycle. Nature 3: 13–22.
Murgue B, Murri S, Triki H, Deubel V, Zeller HG (2001). West Nile in the Mediterranean basin: 1950–2000. Ann. N. Y. Acad. Sci. 951: 117–126.
http://dx.doi.org/10.1111/j.1749-6632.2001.tb02690.x
PMid:11797769
Ng T, Hathaway D, Jennings N, Champ D, Chiang YW, Chu HJ (2003). Equine vaccine for West Nile virus. Dev. Biol. 114: 221– 227.
Nguyen BT, Koh G, Lim HS, Chua AJ, Ng MM, Toh CS (2009) Membrane–based electrochemical nanobiosensor for the detection of virus. Anal. Chem. 81: 7226–7234.
http://dx.doi.org/10.1021/ac900761a
PMid:19663392
Papin JF, Vahrson W, Dittmer DP (2004). SYBR green–based real–time quantitative PCR assay for detection of West Nile Virus circumvents false–negative results due to strain variability. J. Clin. Microbiol. 42: 1511–1518.
http://dx.doi.org/10.1128/JCM.42.4.1511-1518.2004
PMid:15070997 PMCid:PMC387603
Paramasivan R, Mishra AC, Mourya DT (2003). West Nile virus: the Indian scenario. Indian J. Med. Res. 118: 101–108.
PMid:14700342
Parida M, Posadas G, Inoue S, Hasebe F, Morita K (2004). Real–time reverse transcription loop–mediated isothermal amplification for rapid detection of West Nile virus. J. Clin. Microbiol. 42: 257–263.
http://dx.doi.org/10.1128/JCM.42.1.257-263.2004
PMid:14715762 PMCid:PMC321710
Pattan SR, Dighe NS, Bhawar SB, Gaware VM, Hole MB, Musmade DS, Bhosale MS (2009). West Nile Fever: An Overview. J. Biomed. Sci. and Res. 1: 33–48.
Paul SD, Rajagopalan PK and Sreenivasan MA (1970). Isolation of the West Nile virus from the frugivorous bat, Rousettus leschenaulti. Indian J. Med. Res. 58: 1169–1171. PMid:5534069
Pfeffer M, Dobler G (2010). Emergence of zoonotic arboviruses by animal trade and migration. Parasit. Vectors 3: 35.
http://dx.doi.org/10.1186/1756-3305-3-35
PMid:20377873 PMCid:PMC2868497
Platonov AE, Fedorova MV, Karan LS, Shopenskaya TA, Platonova OV, Zhuravlev VI (2008). Epidemiology of West Nile infection in Volgograd, Russia, in relation to climate change and mosquito (Diptera: Culicidae) bionomics. Parasitol. Res.103: 45–53.
http://dx.doi.org/10.1007/s00436-008-1050-0
PMid:19030885
Ratho RK, Sethi S, Prasad SR (1999). Prevalence of Japanese encephalitis and West Nile viral infections in pig population in and around Chandigarh. J. Commun. Dis. 31: 113–116. PMid:10810598
Reisen WK, Fang Y, Martinez VM (2006). Vector competence of Culiseta incidens and Culex thriambus for West Nile virus. J. Am. Mosq. Control Assoc. 22: 662– 665.
http://dx.doi.org/10.2987/8756-971X(2006)22[662:VCOCIA]2.0.CO;2
Rey FA, Heinz FX, Mandl C, Kunz C, Harrison SC (1995). The envelope glycoprotein from tick–borne encephalitis virus at 2 Angstrom resolution. Nature 375: 291–298.
http://dx.doi.org/10.1038/375291a0
PMid:7753193
Richards SL, Anderson SL, Lord CC, Smartt CT, Tabachnick WJ (2012). Relationships between infection, dissemination, and transmission of West Nile virus RNA in Culex pipiens quinquefasciatus (Diptera: Culicidae). J. Med. Entomol. 49: 132–142.
http://dx.doi.org/10.1603/ME10280
PMid:22308781 PMCid:PMC3320798
Risbud AR, Sharma V, Mohan Rao CVR, Rodrigues FM, Shaikh BH, Pinto BD, Verma SP (1991). Post–epidemic serological survey for JE virus antibodies in South Arcot district (Tamil Nadu). Indian J. Med. Res. 93: 1–5.
PMid:1850713
Rodrigues FM, Guttikar SN, Pinto BD (1981). Prevalence of antibodies to Japanese encephalitis and West Nile viruses among wild birds in the Krishna– Godavari Delta, Andhra Pradesh, India. Trans. R. Soc. Trop. Med. Hyg. 75: 258–262.
http://dx.doi.org/10.1016/0035-9203(81)90330-8
Rodrigues FM, Singh BP, Dandawate CN, Soman RS, Guttikar SN, Kaul HN (1980). Isolation of Japanese encephalitis and West Nile viruses from mosquitoes collected in Andhra Pradesh. Indian J. Parasitol. 4: 149–153.
Roehrig JT (2013). West Nile virus in the United States — A Historical Perspective. Viruses 5: 3088–3108.
http://dx.doi.org/10.3390/v5123088
PMid:24335779 PMCid:PMC3967162
Roehrig JT, Nash D, Maldin B, Labowitz A, Martin DA, Lanciotti RS, Campbell GL (2003). Persistence of virus–reactive serum immunoglobulin m antibody in confirmed west nile virus encephalitis cases. Emerg. Infect. Dis. 9: 376–379.
http://dx.doi.org/10.3201/eid0903.020531
PMid:12643836 PMCid:PMC2958550
Rondini S, Pingle MR, Das S, Tesh R, Rundell MS, Hom J, Stramer S, Turner K, Rossmann SN, Lanciotti R, Spie EG, Mu-oz–Jordán J, Larone D, Spitzer E, Barany F, Golightly LM (2008). Development of Multiplex PCR–Ligase Detection Reaction Assay for Detection of West Nile Virus. J. Clin. Microbiol. 46: 2269–2279.
http://dx.doi.org/10.1128/JCM.02335-07
PMid:18495862 PMCid:PMC2446923
Rossi SL, Ross TM, Evans JD (2010). West Nile virus. Clin. Lab. Med. 30: 47–65.
http://dx.doi.org/10.1016/j.cll.2009.10.006
PMid:20513541 PMCid:PMC2905782
Sampatkumar P (2003). West Nile Virus: Epidemiology, Clinical Presentation, Diagnosis, and Prevention. Mayo. Clin. Proc. 78: 1137–1144.
http://dx.doi.org/10.4065/78.9.1137
Shi PY, Kramer LD (2003). Molecular detection of West Nile virus RNA. Expert. Rev. Mol. Diagn. 3: 357–366.
http://dx.doi.org/10.1586/14737159.3.3.357
PMid:12779009
Shukla J, Saxena D, Rathinam S, Lalitha P, Joseph CR, Sharma S, Soni M,Rao PVL, Parida M (2012). Molecular detection and characterization of West Nile virus associated with multifocal retinitis in patients from southern India. Int. J. Infect. Dis. 16: 53–59.
http://dx.doi.org/10.1016/j.ijid.2011.09.020
PMid:22099888
Smithburn KC, Hughes TP, Burke AW, Paul JH (1940). A neurotropic virus isolated from the blood of a native of Uganda. Am. J. Trop. Med. 20: 471–492.
Suthar MS, Diamond MS, Gale M Jr (2013). West Nile virus infection and immunity. Nat. Rev. Microbiol. 11: 115.
http://dx.doi.org/10.1038/nrmicro2950
PMid:23321534
Vanlandingham DL, McGee CE, Klinger KA, Vessey N, Fredregillo C, Higgs S (2007). Relative susceptibilties of South Texas mosquitoes to infection with West Nile virus. Am. J. Trop. Med. Hyg. 77: 925–928.
PMid:17984355
Varma MGR (1960). Preliminary studies on the infection of Culicine mosquitoes with the Tamil Nadu strain of West Nile virus. Ind. J. Med. Res. 48: 537–548.
Vazquez A, Sanchez–Seco MP, Ruiz S, Molero F, Hernandez L, Moreno J. (2010). Putative new lineage of West Nile virus, Spain. Emerg. Infect. Dis. 16: 549–552.
http://dx.doi.org/10.3201/eid1603.091033
PMid:20202444 PMCid:PMC3322021
Westaway EG, Mackenzie JM, Khromykh AA (2002). Replication and gene function in Kunjin virus. Curr. Top. Microbiol. Immunol. 267: 323–351.
http://dx.doi.org/10.1007/978-3-642-59403-8_16
PMid:12082996
Vitek CJ, Richards SL, Mores CN, Day JF, Lord CC (2008). Arbovirus transmission by Culex nigripalpus in Florida. J. Med. Entomol. 45: 483– 493.
http://dx.doi.org/10.1603/0022-2585(2008)45[483:ATBCNI]2.0.CO;2



