Advances in Animal and Veterinary Sciences
Research Article
Advances in Animal and Veterinary Sciences 2 (7): 393 – 400Interaction of ß – Benzenehexachloride and Cadmium on Physiologic Response of Laying Domestic Mallard (Anas Platyrhynchos L.)
Renato S.A. Vega1*, Devita Anggraeni2, Henry I. Rivero3, Rainie Rich Chucky S. Yambao1, Marie Fe M. Iranzo1, Carla Alilie A. Lontoc1, Severino S. Capitan1
- Animal Breeding & Physiology Division, ADSC, College of Agriculture, U.P. Los Baños, College, Laguna, PHILIPPINES 4031
- Surgery & Radiology Division, Faculty of Veterinary Medicine, Gadjah Mada University, INDONESIA
- MSU–Iligan Institute of Technology, A. Bonifacio Ave., Iligan City 9200 Philippines
*Corresponding author:rsvega@up.edu.ph
ARTICLE CITATION:
Vega RSA, Anggraeni D, Rivero HI, Yambao RRCS, Iranzo MFM, Lontoc CAA, Capitan SS (2014). Interaction of ß – benzenehexachloride and cadmium on physiologic response of laying domestic mallard (anas platyrhynchos L.). Adv. Anim. Vet. Sci. 2 (7): 393 – 400.
Received: 2014–05–01, Revised: 2014–08–03, Accepted: 2014–08–05
The electronic version of this article is the complete one and can be found online at
(
http://dx.doi.org/10.14737/journal.aavs/2014/2.7.393.400
)
which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited
ABSTRACT
This study aims to understand the effects of cadmium and β–Benzenehexachloride (β–BHC) administered in feed on the physiological response of laying Mallard. Laying ducks (n=120) at three different stages (I: 33–37, II: 38–42 and III: 43–47 weeks old) were randomly allotted into 3x2 factorial design in CRD with 3 Cd and 2 BHC doses, as first and second factor respectively replicated in 4. The doses were doubled (II) and tripled (III) from 0.017 (Cd1), 0.034 ppm Cd (Cd2), and 0.033 ppm β–BHC (P1) of stage I. Hematology, hepatosomatic index (HSI), gonadosomatic index (GSI) and oviductosomatic index (OVI) were compared. At stage I, Cd and β–BHC did not have any significant effects at all parameters observed. Significant effects were observed only at stage II and III. The HSI increased and decreased with 0.033 and 0.067 of Cd treatment, respectively, however β–BHC increased HSI at stages II and III (P<0.05). The OVI was reduced with 0.067Cd, but in combination with 0.067β–BHC it was enlarged (P<0.05). The number of RBC was significantly reduced by β–BHC. The number of WBC was reduced by Cd levels but β–BHC elevated it by 20x103/µl (P<0.05). Percent heterophil was suppressed by Cd but elevated by β–BHC (P<0.05). Lymphocyte was suppressed by 0.1 ppm β–BHC, while monocyte was enhanced by the levels of Cd, but in combination with β–BHC it was reduced (P<0.05). Briefly, HSI, OVI, WBC and heterophil were suppressed by Cd, and lymphocyte and RBC were suppressed by β–BHC. Therefore, individual and low levels of Cd and β–BHC seem tolerable, but in combination their effects are compounded and threaten productivity and food safety. Over–all it is suggested that the combined tolerable limits of heavy metals and pesticide residue should be established for future research and regulatory purposes.
INTRODUCTION
The world’s duck egg production is dominated by the Asian countries, probably because it provides low feed cost, feeds not competing to man and eggs providing daily source of income. Apparently waterfowls are considered good sources of healthy meat and eggs. The Philippines ranks number 5 in Mallard production, but slowly declining over the years due to lack of breeding program, rising feed cost, low price of eggs and the presence of pollutants in feeds are unknowingly affecting productivity (Vega, 2009, Vega et al., 2011, Anggraeni et al., 2012).
The organochlorine pesticides (OCPs) were found to posses multiple health risks and their presence in high amounts has become a major environmental and health concern (Aktar et al., 2009). Among these OCPs is β–Benzenehexachloride (β–BHC). Kojima et al., (2004) studied 200 possible endocrine disrupting properties of 200 pesticides and among those in the list of organochlorines were o,p´–Dichorodiethyltrichloroethane (o,p'–DDT), β–BHC, methoxychlor, and α–endosulfan. In ovo studies showed that β–BHC can negatively affect growth and development of ducks (Vega et al., 2014). Upon the discovery of the environmental and health hazards of pesticides, specifically OCPs, the use of these compounds is now being regulated.
Another substance that is also currently of concern is cadmium (Cd), which is naturally present in the soil and mined minerals used in agricultural production such are the phosphorus fertilizers for crops and phosphate supplements for feeds. According to Klasing et al. (2005), Cd tends to accumulate in the kidney, liver, pancreas and spleen. However, Cd is not easily transported into eggs and milk (Klasing, et al., 2005; Vega et al., 2014). The effects of Cd in the physiology of farm animals and humans alike are still being studied. In chickens, Cd is said to be toxic to immune cells possibly by inducing oxidative stress resulting to DNA damage and apoptosis (Li et al., 2010). Previous research by Cain et al., (1983) in Mallard ducks reported a significant decrease in Packed Cell Volume (PCV) and Hemoglobin (Hb), an increase in glutamic pyruvic transaminase (GPT) and the presence of kidney damage upon treatment with 20 ppm Cd. They also observed Cd accumulation in the liver. The separate studies of Vega et al., (2011) and Anggraeni et al., (2012) were in agreement that Cd can cause damage and decrease in liver size; and enhance early laying days by 35 days in ducks. Also, along with organochlorines such as β–BHC, γ–Benzenehexachloride and γ–chlordane, Cd causes liver damage.
Productivity and food safety are the main concerns in determining the effects of these unwanted residues in feeds. Thus, this study aimed to investigate the individual and the interacting effects of β–Benzenehexachloride (β–BHC) and Cadmium (Cd) in feed on physiologic and haematologic responses of domestic Mallard Ducks at sexual maturity to support the previously reported detrimental effects of these compounds in Mallard ducks (Cain et al., 1983; Vega, 2009; Vega et al., 2011; Anggraeni et al., 2012; Vega et al., 2014).
MATERIALS AND METHODS
Pre–Experimental Conditions of Mallard
Female ducks used in this study were supplied by a single farm in Cavite, samples of female (n=10) and male (n=10) ducks at 16 weeks of age were sacrificed one day after the arrival for the extraction and isolation of blood, liver and gonad samples to have an initial assessment of the Mallards.
The specimens or samples for pesticide residue and cadmium analysis were taken by pooling liver samples. Liver samples were collected and pooled for female and male ducks, because they originate from the same sources having common feed and environment. All the samples were submitted to Natural Sciences Research Institute (NSRI; PNS ISO/IEC 17025:2005) of the University of the Philippines, Diliman, Quezon City for pesticide residue and cadmium analysis. The same assay protocol for Cd and Organochlorine Pesticide Residues (OCPs) was used as described elsewhere (Vega et al., 2014)
Treatment Administration
After four months of growing or adjustment period, the 33 weeks old laying ducks (n = 120) for three different stages was accomplished with four weeks interval for every stage and randomly allotted into 3 x 2 Factorial design in CRD (3 levels of Cd: 2 levels of β–BHC). The levels in Stage I were; (1) Cd0P0 = 0Cd+0BHC, (2) Cd0P1 = 0Cd+0.033BHC, (3) Cd1P0 = 0.017Cd+0BHC, (4) Cd1P1 = 0.017Cd+0.033BHC, (5) Cd2P0 = 0.034Cd+0BHC, (6) Cd2P1 = 0.034Cd+0.033BHC. The level/dose of six treatments in stage I was doubled and tripled for stage II and III, respectively.
Pure grade CdCl2 and β – BHC were procured from Supelco (Indonesia). The residue dosages set was the amount found in feed, these were 0.05 and 0.10 ppm Cd; and 0.10 ppm β – BHC. These were gradually obtained in stage III. Acetone (Analytical Grade, Scharlau) was used as vehicle for all the contaminants for inclusion in the commercial feed. Amount of feed given to the duck was 130 g duck–1day–1. Each treatment was replicated 4 times with 5 ducks per replicate. The experimental period lasted for 14 weeks. Treatments were given to laying ducks at stage I, II, and III, i.e. 33–37; 38–42 and 43–47 weeks of age, respectively. The sampling procedure were done on the last day of different stages, one duck per replicate were sacrificed. The HSI, OVI and GSI were measure in three different stages (I, II and III) while the haematological changes were determined in stage I and stage III only for comparison.
The data were analyzed using SAS version 6.12 using PROC ANOVA having 3x2 factorial experimental design in Complete Randomized Design (CRD). The comparison of treatments means after the ANOVA declared significance at P≤0.05 used LSD. Interaction or two–way table was used when the ANOVA declares significant interaction.
Hematological Response
The hematological response of the ducks at 33 to 47 weeks of age to β–BHC and cadmium was based on the value of packed cell volume, hemoglobin, total red blood cell, total white blood cell and differential leucocyte.
Packed cell Volume (PCV)
Determination of PCV in birds is essentially the same as that for other species. Blood was collected and placed into tubes containing ethylenediamine tetraacetate (EDTA). Tubes were centrifuged at 12,000 x g for 5 minutes and read on a standard microhematocrit graphic reader (Pierson, 2000).
Hemoglobin Concentration
Methods and formula for determining the hemoglobin concentration are identical to those used for mammalian species. In the case of avian hemoglobin determination, samples were centrifuged at 1000 x g for 10 minutes following red cell lysis to remove nuclei and other cellular debris (Pierson, 2000).
Total Red Blood Cell (TRBC)
According to the Natt and Herrick method, blood containing EDTA was drawn into a standard blood dilution pipette to the 0.5 mark, followed by methyl violet 2B diluents to the 101 mark. This gave a final dilution 1:200. The pipette was then briefly agitated, blotted and the counting chamber of a Neubauer hemocytometer charged. After a few minutes to allow settling, the red cell count was determined under high–dry magnification (40x). Erythrocytes appearing in the center and four corner squares were counted and the total multiplied by 1 x 104 to give the TRBC count per microliter (Pierson, 2000).
Total White Blood Cell (TWBC)
The Natt and Herrick method was a direct TWBC determination. For the TWBC count, leucocytes (dark to light blue) appearing in the nine large squares of an improved Neubauer hemocytometer were totaled. The TWBC count was then calculated using the following formula:
TWBC/µl = |
(number of leucocytes in 9 squares) |
x 220 (Pierson, 2000) |
Differential Leucocyte
Differential white blood cell counts require the preparation of stained blood smears. Blood treated with EDTA was used. A drop of blood was placed on a precleaned microscope slide. While the slide was held between the fingers, a rectangular coverglass was placed on top of the blood. As the blood begin to spread, the coverslip was gently slipped off the microscope slide in a horizontal fashion. After air drying, Giemsa stain was used to prepare slides for examination (Pierson, 2000).
Reproductive Response
Reproductive response of the ducks at 33 to 47 weeks of age to organochlorine pesticide and cadmium was based on organosomatic indices (hepatosomatic, gonadosomatic and oviductosomatic). The liver, ovary and oviduct of each duck was carefully removed and placed on aluminium foil for documentation. Total fresh weight of the liver, ovary and oviduct was obtained.
Hepatosomatic Index
Hepatosomatic index (HSI) value was obtained from the ratio of fresh liver weight to live body weight and multiplied by 100. The value was expressed in % of the body weight. Hepatosomatic index is an indicator of hepatic growth or development according to the age and physiological status.
Gonadosomatic Index
The value of gonadosomatic index (GSI) was obtained from the ratio of fresh ovary weight to live body weight and multiplied by100. The value was expressed in % of the body weight.
Oviductosomatic Index
The same as HSI and GSI, oviductosomatic index (OVI) was the ratio of oviduct weight to live body weight and multiplied by 100. The value was expressed in % of the body weight. Oviductosomatic index also can be used to estimate the status of reproductive system.
RESULTS AND DISCUSSIONS
The result of pesticide analysis in commercial feed and duck liver (female and male) are shown in Table 1. Endosulfan II and p,p'–DDD + o,p'–DDT were found in commercial feed with concentrations of 21 and 0.75 ng/g, respectively. In female duck liver, endosulfan II and p,p'–DDD + o,p'–DDT were observed at concentrations of 240 and 0.93 ng/g, respectively. The analysis also revealed the presence of same chemicals (endosulfan II, 200 ng/g and p,p'–DDD + o,p'–DDT, 1.43 ng/g) plus endosulfan sulfate, 6.3 ng/g, in the liver of male ducks. It was reported that β – , γ –, α – BHC were also present in duck commercial feed as reported at levels of 0.088–0.15ppm, 0.08–0.09 ppm, 0.02–0.55 ppm respectively (Vega et al., 2011). The result of cadmium analysis in commercial feed and duck liver (female and male) are shown in Table 2. Cd was present in commercial feed, female and male duck liver at concentrations of 0.049 mg/kg, 0.18 mg/kg and 0.25 mg/kg, respectively. According to AMAP (2004), the maximum permissible concentration of Cd in meat and poultry is 0.05 mg/kg, while provisional tolerable weekly intake (PWTI) of Cd per person is 7 µg/kg body weight (WHO, 2006). Cd was found to exceed the maximum permissible concentration. The presence of pesticides and Cd in the liver was presumably due to the ranging or foraging system before they were purchased.
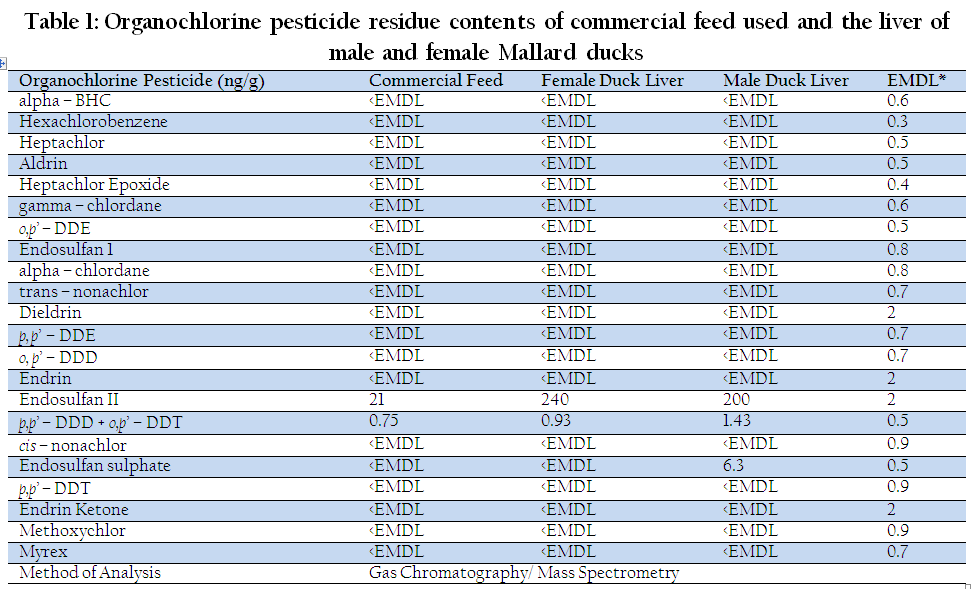
Table 1: Organochlorine pesticide residue contents of commercial feed used and the liver of male and female Mallard ducks
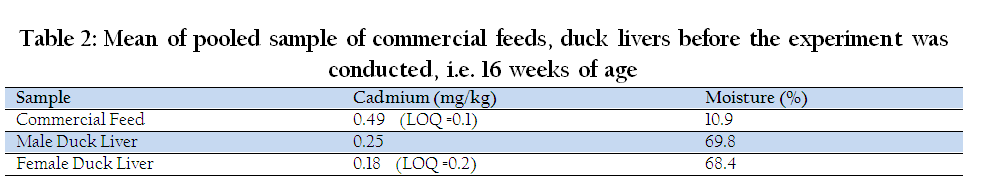
Table 2: Mean of pooled sample of commercial feeds, duck livers before the experiment was conducted, i.e. 16 weeks of age
Organosomatic indices (HSI, GSI and OVI) were measured when the ducks were at 33 weeks of age, i.e. before the treatment were administered. The results showed that the means of HSI, GSI and OVI were 3.07 ± 0.16, 2.93 ± 1.5 and 2.91 ± 1.34, respectively, which is comparable to those reported elsewhere (Rivero et al., 2009) Cd and β – BHC did not affect HSI within stage I. But, there was significant effect of Cd on HSI at stage II (P<0.05) and at low concentration of Cd (0.034 ppm), significantly higher HSI than the control was observed (Table 3). Significant effect of β – BHC on HSI was also observed at stage II and stage III (P<0.05). At stage II, β – BHC (0.067 ppm) significantly increased HSI, and, increasing concentration of β – BHC (0.10 ppm) similarly increased HSI from stage II to III (P<0.05). The present study showed that higher treatment with Cd increased the liver weight. This result was corroborated by Demerdash et al., (2004) who reported that treatment with CdCl2 (5 mg/kg BW) in male rat caused an increase in the weight of liver. Cadmium and organochlorine pesticide were considered as EDCs which have the ability to mimic the action of estrogen. The study conducted by Schubert et al., (2008) showed that hepatosomatic index (HSI) was significantly increased in brown male trout exposed to mixtures of estrone, 17β– estradiol and 4–nonylphenol for 150 days until the onset of spawning, which indicated an enhanced liver metabolism due to the increased VTG synthesis in the liver. There was no significant interaction effect of Cd and β – BHC on HSI at all stages. However, significant differences were observed on HSI at stage II (P<0.05). The highest mean HSI was noted in 0.033 ppm Cd + 0.067 ppm β–BHC (3.27%) this was significantly higher than control (2.68%) and 0.067ppm Cd alone (2.67%). This indicates that combination of Cd (0.033 ppm) and β–BHC (0.067 ppm) were more potent in activating HSI compared to the control and Cd (0.067 ppm) alone.
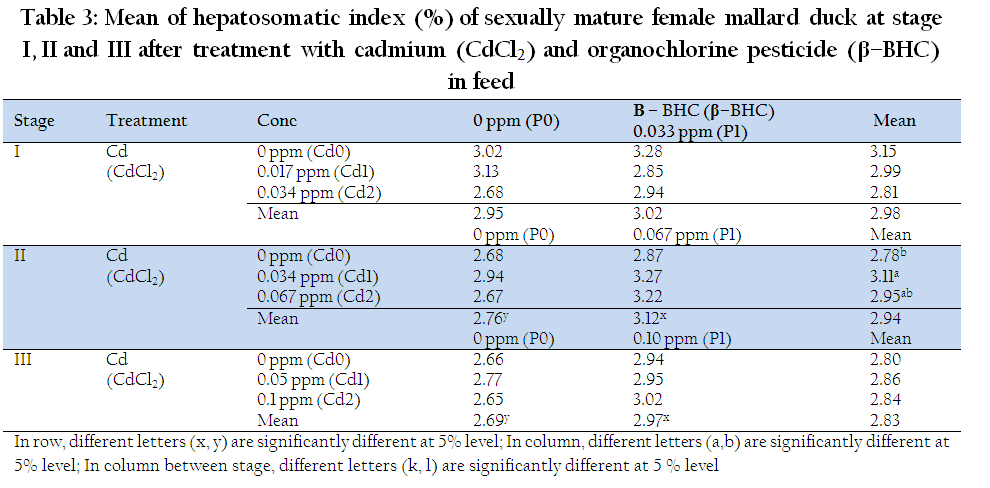
Table 3: Mean of hepatosomatic index (%) of sexually mature female mallard duck at stage I, II and III after treatment with cadmium (CdCl2) and organochlorine pesticide (β–BHC) in feed
Individual Cd and β–BHC residue did not affect OVI at stage I, II, and III. However, there was significant interaction effect of Cd and β–BHC on OVI at stage I and II (P<0.05; Table 4). Combination of Cd+β–BHC, revealed significantly lowest OVI (1.76 %), this was even lower than 0.034ppm Cd (3.04 %) +β–BHC indicative of combined downsizing action of Cd at lower levels. Longer exposure and doubly higher dose of Cd and BHC (Stage II), revealed that OVI has tendency to become smaller at lower dose of Cd (0.034ppm) than at higher dose of Cd (0.067ppm). Interestingly, 0.067Cd+0.067BHC significantly increased the OVI (3.65 %), an observation showing a clear manifestation that combined Cd and β–BHC at longer and higher doses have direct influence on hormonal response in female Mallards, being able to mimic the activity of estrogen as xenoestrogens.
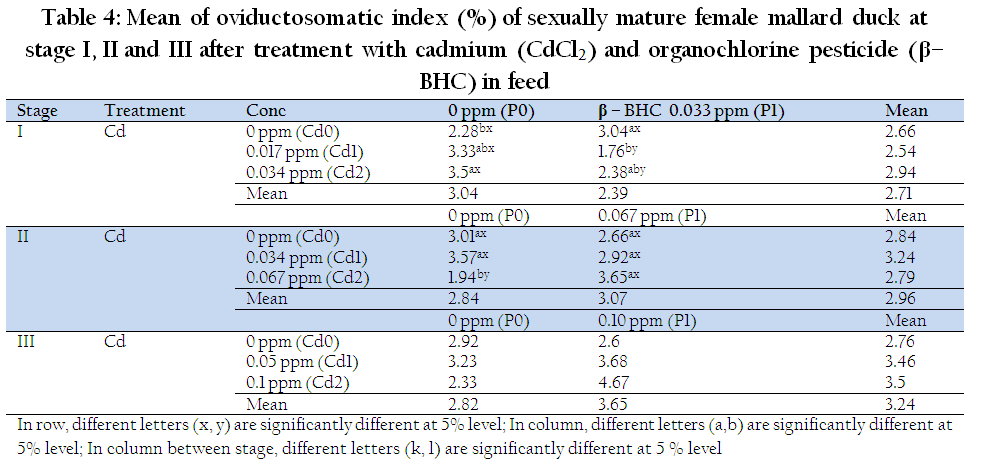
Table 4: Mean of oviductosomatic index (%) of sexually mature female mallard duck at stage I, II and III after treatment with cadmium (CdCl2) and organochlorine pesticide (β–BHC) in feed
The nonsignificant effect on OVI at last stage (III) of combined and highest desired level of Cd and β–BHC is suggestive of ducks developing resistance or tolerance to these residues for three months of exposure. The study conducted by Wade et al., (2002) showed that exposure to low or moderate doses of a mixture of persistent organochlorines and metals affects liver physiology and has some effects on the function of T cells in the adult male rats. Kohler et al., (1969) as cited by Berg et al., (2001) stated that estrogen is the prerequisite for differentiation and development of the oviducts in female birds. Proliferation of the luminal epithelium and differentiation of ciliated and glandular cells are estrogen–controlled processes. Therefore, increasing weight of the oviduct as shown in this study might be due to the effect of Cd and β–BHC in mimicking the action of estrogen.
The control treatment values for the PCV, RBC and WBC were 40.9%, 3.24 (x106/µl) and 16.6 (x103/µl), respectively. Meanwhile, the values for the percent heterophil, lymphocyte and monocyte were 33.6, 59.6 and 5.4% respectively. The several reported values of hematology for normal duck and chicken (Campbell, 2000; Okeudo et al., 2003; Orawan and Aengwanich, 2007) revealed that our values were within the normal range.
The individual effect of β–BHC and Cd on PCV showed no significant differences at stage I and III. Combined Cd and β–BHC did not affect PCV at stage I (data not shown). There was significant interaction effect of Cd and β–BHC on PCV at stage III (Table 5.). The combined effects (0.1Cd + 0.1BHC), significantly decreased PCV (36.25 %). This observed decrease in PCV by combined Cd and β–BHC was similar to the decrease in PCV as reported by Cain et al., (1983) in 8 week–old ducklings exposed to 20 ppm Cd (200x higher). Hence, low levels but combined effect Cd and β – BHC may have a compounding action as endocrine disruptors. It is possible that increasing vitellogenin (VTG) may have resulted to the decreasing level of PCV. According to William et al., (2004), PCV may decline because of an increase in blood volume (hemodilution), which is an osmotic adjustment in response to the appearence of substantial levels of VTG in the plasma.
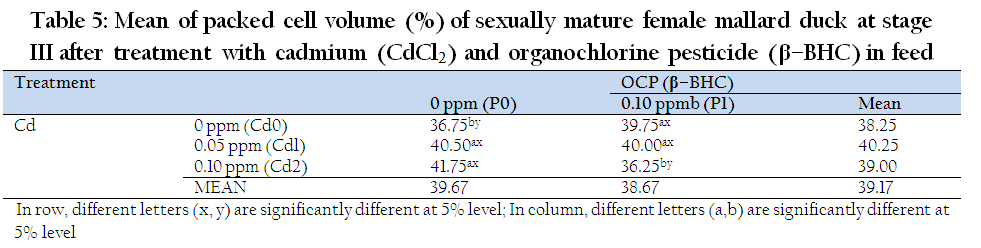
Table 5: Mean of packed cell volume (%) of sexually mature female mallard duck at stage III after treatment with cadmium (CdCl2) and organochlorine pesticide (β–BHC) in feed
Cadmium and β–BHC significantly affect WBC at stage III (Figure 1). Increasing concentration of Cd (0.05 ppm and 0.1 ppm) significantly decreased WBC, on the other hand, β–BHC (0.10 ppm) significantly increased WBC. The the effect of low level of Cd (0.05 to 0.10 ppm) in our study is contrary to the 5 ppm Cd study conducted in rats by Demerdas et al., (2004). However, in fish (Ghiasi et al., 2010) the decreased WBC of Cd exposed fish suggested an immunological suppression. Note that oviparous and mammals may have differing sensitivity. In oviparous animals, higher level lindane (5 ppm) immuno suppression and stress effect were also observed after adiministration twice in a week in 6 species of agriculturally important birds (Mandal et al., 1986). Noticeably our experiment of lower level Cd doses significantly reduced number of WBC, hence immune response sensitivity.
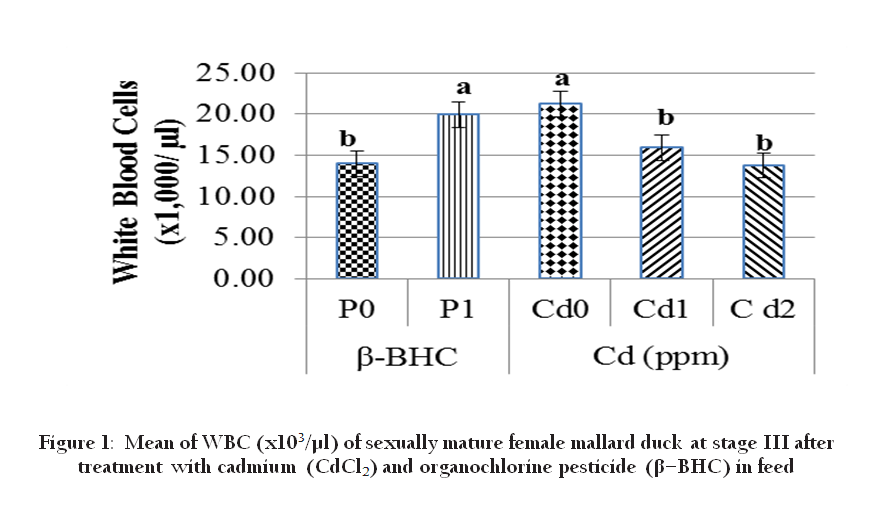
Figure 1: Mean of WBC (x103/µl) of sexually mature female mallard duck at stage III after treatment with cadmium (CdCl2) and organochlorine pesticide (β–BHC) in feed
Organochlorine pesticides are known to have the ability to induce oxidative stress according to Vrbanac et al., (2002). The number of RBC, % heterophils, % lymphocytes, and % monocytes are stress–affected haematological responses. Thus the number of RBC at stage III was reduced significantly by 0.1ppm BHC (Figure 2). Normal RBC function depends on the integrity of cell membrane which is the target of many toxic agents including pesticide (Banerjee et al., 1999). Organochlorine pesticide is known to have the ability to induce oxidative stress according to Vrbanac et al., (2002). Since the life span of avian RBC is shorter, the body temperature of avian is higher and their metabolism is more intense compared to mammals, therefore, avian metabolism will need more oxygen when they are exposed to a higher oxidative stress (Vrbanac et al., 2002). Thus, there is a possibility that RBC is compensated by increasing the number from stage I to III to get more oxygen for metabolism.
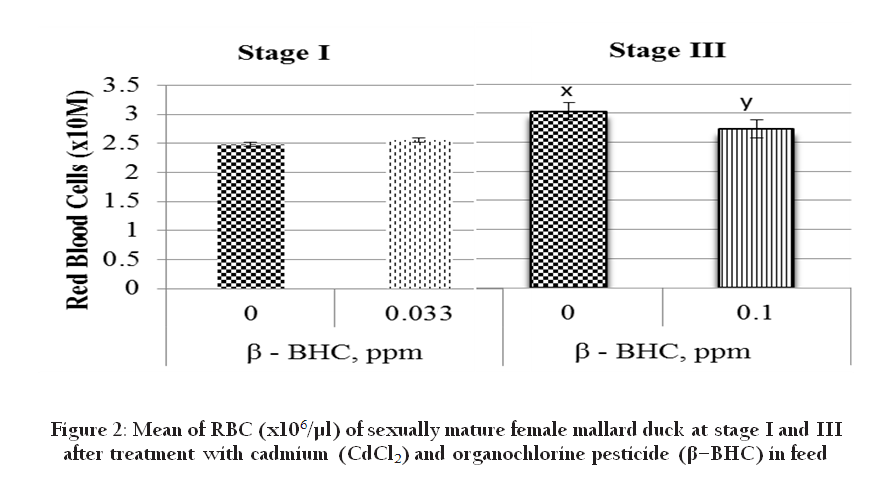
Figure 2: Mean of RBC (x106/µl) of sexually mature female mallard duck at stage I and III after treatment with cadmium (CdCl2) and organochlorine pesticide (β–BHC) in feed
The % heterophil on the other hand was increased in combination with 0.1ppm BHC at stage III (Table 6), Cd might have activated the phagocytic action of heterophil but with BHC the higher concentration indicates inactivation of phagocytic action of heterophil (Davis et al., 2008). Phagocytosis may cause excessive active oxygen in organism and intensifies the active oxygen action on the lipid peroxidation and results in oxidation injury. Our results showed that the combination of Cd (0.05 ppm) and β–BHC (0.10 ppm) were more potent in increasing heterophil compared to Cd (0.05 ppm and 0.1 ppm) alone as observed in our study

Table 6: Mean of heterophil (%) of sexually mature female mallard duck at stage III after treatment with cadmium (CdCl2) and organochlorine pesticide (β–BHC) in feed
At stage III, there was no significant effect of Cd on lymphocyte (Figure 3), however β–BHC (0.1ppm) significantly decreased lymphocyte and residue exposure with stages, i.e. stage I having higher lymphocyte than III. Since lymphocyte is associated to stress, the lowering across the stage may be due stress–related exposure to Cd and β – BHC. Altered lymphocyte proliferation was significantly associated with elevated blood levels of organochlorines (Finkelstein et al., 2007). According to Grasman et al., (1996), organochlorines are known to be positively associated with avian immune response. It is known that organochlorine can stimulate stress effect in birds (Mandal et al., 1986). Stress stimulates reduction in circulating lymphocyte numbers due to glucocorticoid–induced alterations in the redistribution of lymphocyte from the blood to other body compartments (Davis et al., 2008). Monocytes are phagocytic cells which are associated with defence against infections and bacteria (Davis et al., 2008). There was no significant individual effect of Cd and β–BHC on monocyte at stage III, but highly significant interaction was observed (P<0.01). At stage I, lowest dose of β–BHC (0.033 ppm) significantly increased monocyte (6.5 vs 2.34), but not Cd. Monocyte of Control (Cd0P0; 2.25 %) was not significantly different with 0.1 ppm β–BHC (Cd0P1) and 0.05 ppm Cd (Cd1P0). Clearly with the increasing level of Cd, monocyte increased, but in combination with 0.1 ppm β–BHC, monocyte decline proportionally (Table 7). According to Lall and Dan (1999), cadmium at a very high dosage (30, 100 and 300 ppm) in mice suppressed both humoral and cellular immunity with simultaneous increase in the level of blood corticosterone. Increase of corticosterone concentrations due to stress may reduce monocyte accumulation at inflammatory sites. Apparently in our studies, even if the level of Cd is very low (<0.10 ppm) if it is combined with β–BHC, the impact on stress is similar to high Cd level as manifested by the decline in monocyte. This corticosterone stimulated reduction in macrophage numbers can also be attributed to inhibition of enzymes for phagocytosis and inhibition of secretion of interleukin–2, the cytokine needed for proliferation and maturation of lymphocytes in chickens (Mumma et al., 2006).
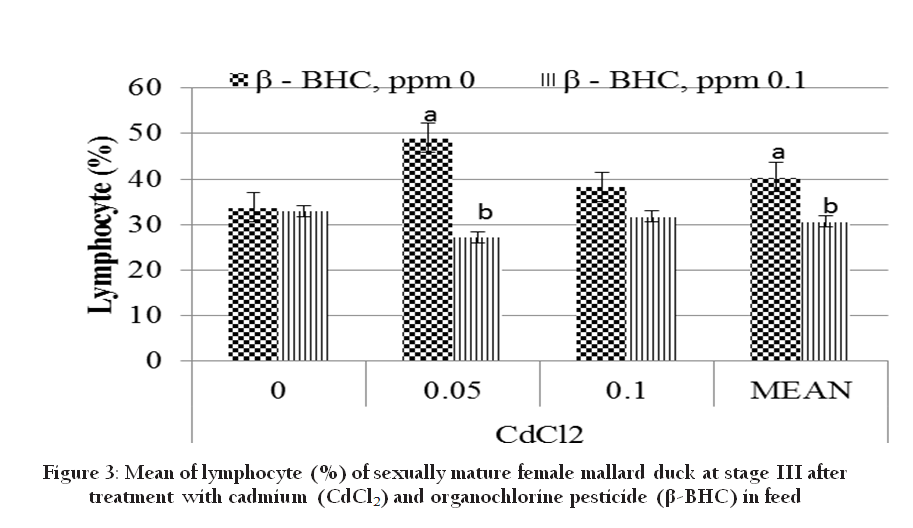
Figure 3: Mean of lymphocyte (%) of sexually mature female mallard duck at stage III after treatment with cadmium (CdCl2) and organochlorine pesticide (β-BHC) in feed

Table 7: Mean of monocyte (%) of sexually mature female mallard duck at stage III after treatment with cadmium (CdCl2) and organochlorine pesticide (β–BHC) in feed
Briefly, Hepatosomatic Index, Ovarysomatic Index, number of White Blood Cells and % heterophil were suppressed by Cd, while β – BHC suppressed the % lymphocyte and number of Red Blood Cells. The combination of both residues reduced percent Packed Cell Volume and % monocyte. Apparently, Cd is more a productivity and food safety threat than β–BHC as Cd suppressed HSI, OVI and many other important haematological response parameters. However even if the Cd level is low (<0.10 ppm), the negative effect is enhanced by BHC and the phagocytic activation done by Cd is inactivated by the organochlorine pestice residue or β–BHC. Interaction effects of both Cd and BHC masked their individual effects on haematological parameters, and their combined effects even at low levels are compounding hence becoming a silent threat to domestic Mallard productivity.
ACKNOWLEDGEMENT
The authors would like to express their gratitude to the UP Systems Emerging Science and Technology Grant for providing the equipment, first year budget operation by the Department of Agriculture–Bureau of Agricultural Research (DA–BAR) and the third year budget for operation by the DA–Biotech Program Implementation Unit (DA–Biotech PIU), headed by Drs. Amelia Guevara, Nicomedes Eleazar and Candida B. Adalla, respectively. Special thanks are also extended to Moody, Cenon, Jun and Bido, the staff of the Animal Breeding and Physiology Division, for their helping hands and services during weekends and holidays.
REFERENCES
Aktar, MW, Sengupta D, Chowdhury A (2009). Impact of pesticides use in agriculture: their benefits and hazards. Interdisc. Toxicol. 2(1): 1 – 12.
http://dx.doi.org/10.2478/v10102-009-0001-7
PMid:21217838 PMCid:PMC2984095
AMAP (Arctic Monitoring Assessment Programme) (2004). PTS limits and levels of concern in the environment, food and human tissues. In : Persistent Toxic Substances, Food Security and Indigenous Peoples of the Russian North. Retrieved August 14, 2010, from http://www.amap.no/documents/index.cfm?dirsub=%2FPersistent%20Toxic%20Substances%2C%20Food%20Security%20and%20Indigenous%20Peoples%20of%20the%20Russian%20North
Anggraeni D, Capitan SS, Sevilla CC, Vega RSA, Cruzana BC (2011). Effects of Cadmium and Pesticide on the Liver of Sexually Mature Female Philippine Mallard Duck (Anas platyrhynchos L.). PJVAS Vol. 37(1):67–76.
Banerjee BD, Seth V, Bhattacharya A (1999). Biochemical effects of some pesticides on lipid peroxidation and free radical scavenger. Toxicol. Letters. 107: 33 – 37.
http://dx.doi.org/10.1016/S0378-4274(99)00029-6
Cain BW, Sileo L, Franson JC, Moore J. (1983). Effects of dietary cadmium on mallard ducklings. Environ Res 32(2): 286 – 297.
http://dx.doi.org/10.1016/0013-9351(83)90112-3
Campbell TW (2000). Normal hematology of waterfowl. In : Feldman BF, Zinkl JG and Jain NC. Ed. Schalm's Veterinary Hematology, Lippincott Williams & Wilkins, Philadelphia. pp. 1161–1163.
Klasing, KC, Goff, JP, Greger, JL, King, JC, Lall, SP, Lei, XG, Lin, JG, Nielsen, FH, Spears, JW, Lewis, AJ, Jonker, JS, Jameison, DL, and Tsai, P 2005. Mineral Tolerance of Animals, 2nd ed. National Academies Press. 491 pp., (2005) by the Committee on Minerals and Toxic Substances in Diets and Water for Animals, Board on Agriculture and Natural Resources, Division on Earth and Life Studies, National Academy of Sciences, USA.
Davis AK, Maney DL, Maerz JC (2008). The use of leukocyte profiles to measure stress in vertebrates: a review for ecologists. Functional Ecology. 22: 760 – 772.
http://dx.doi.org/10.1111/j.1365-2435.2008.01467.x
Demerdash FM, Yousef MI, Kedwany FS, Baghdadi HH (2004). Cadmium– induced changes in lipid peroxidation, blood hematology, biochemical parameters, and semen quality of male rats : protective role of vitamin E and β–carotene. Food Chem. Toxicol. 42 : 1563–1571.
http://dx.doi.org/10.1016/j.fct.2004.05.001
PMid:15304303
Finkelstein ME, Grassman KA, Croll DA, Tershy BR, Keitt BS, Jarman WM, Smith DR (2007). Contaminant–associated alteration of immune function in black–footed albatross (Phoebastria nigripes), a north pacific predator. Environ. Toxicol. Chem. 26: 9.
http://dx.doi.org/10.1897/06-505R.1
PMid:17702543
Ghiasi F, Mirzargar SS, Badakhshan H, Shamsi S (2010). Effects of low concentration of cadmium on the level of lysozyme in serum, leucocyte count, and phagocytic index in Cyprinus carpio under the wintering condition. J. Fish. Aquat. Sci. 5: 113 – 119.
http://dx.doi.org/10.3923/jfas.2010.113.119
Kohler P, Grimley PM, O'Malley BW (1969). Estrogen–induced cytodifferentiation of the ovalbumin–secreting glands of the chicks oviduct. In : Berg C, Holm L, Brandt I and Brunstrom B. 2001. Anatomical and Histological Changes in the Oviducts of Japanese Quail, Coturnix japonica, after Embryonic Exposure to Ethynyloestradiol. Reproduction. 121: 155 – 165.
Kojima H, Katsura E, Takeuchi S, Niiyama K, Kobayashi K (2004). Screening for Estrogen and Androgen Receptor Activities in 200 Pesticides by In Vitro Reporter Gene Assays Using Chinese Hamster Ovary Cells. Environ. Health Pers. 112(5):524-531.
http://dx.doi.org/10.1289/ehp.6649
PMid:15064155 PMCid:PMC1241915
Lall SB, Dan G (1999). Role of Corticosteroids In Cadmium Induced Immunotoxicity. Drug Chem. Toxicol. 22(2): 401– 409.
http://dx.doi.org/10.3109/01480549909017843
PMid:10234475
Li JL, Li HX, Li S, Tang ZX, Xu SW, Wang XL (2010). Oxidative Stress–Mediated Cytotoxicity of Cadmium in Chicken Splenic Lymphocytes. Polish J. of Environ. Stud. 19(5): 947 – 956.
Mumma JO, Thaxton JP, Thaxton YV, Dodson WL (2006). Physiological Stress in Laying Hens. Poultry Sci. 85: 761 – 769.
http://dx.doi.org/10.1093/ps/85.4.761
PMid:16615361
Okeudo NJ, Ocoli IC, Igwe GO (2003). Hematological Characteristics of Ducks (Cairina moschata) of Southeastern Nigeria. Tropicultura. 21(2): 61 – 65.
Orawan C, Aengwanich W (2007). Blood cell characteristics, hematological values and average daily gained weight of thai indigenous, thai indigenous crossbred and broiler chicken. Pakistan J. Biol. Sci. 10(2): 302 – 309.
http://dx.doi.org/10.3923/pjbs.2007.302.309
PMid:19070032
Pierson FW (2000). Laboratory techniques for avian hematology. In : Feldman BF, Zinkl JG and Jain NC. Ed. Schalm's Veterinary Hematology, Lippincott Williams & Wilkins, Philadelphia. Pp. 1145–1146.
Rivero HI, Vega RSA, Capitan SS, Lambio AL (2009). Egg–at–First–Lay of Pre–Lay Pateros Ducks Anas platyrhynchos Linn. on Cadmium– and Organochlorine Pesticide Residue–Contaminated Feed Administration. Proceedings of the 2nd International Seminar and 12th Annual National Conference of the Philippine Society for the Study of Nature, 22–27 May 2012, East Asia Royal Hotel, General Santos City, Philippines.
Schubert S, Peter A, Burki R, Schonenberger R, Suter MJF, Segner H, Holm PB (2008). Sensitivity of brown trout reproduction to long–term estrogenic exposure. Aquatic Toxicol. 90: 65 – 72.
http://dx.doi.org/10.1016/j.aquatox.2008.08.002
PMid:18804294
Vega RSA (2009). Effects of endocrine disrupting chemicals in mallard ducks. In SEARCA Professorial Chair Lecture. Retrieved July 10, 2010 from http://www.searca.org/web/adss/2009/handouts/SS_Vega_18Jun2009.pdf.
Vega RSA, Capitan SS, Lambio AL, Garcia BR, Rivero HI (2011). Low Levels of Organochlorine Residues in Combination with Cadmium in Feed at Perpubertal Stage of Domestic Mallard (Anas platyrhynchos Linn.). J. Environ. Sci. Mgt. 14(2): 21 – 27.
Vega RSA, Remoral DMS, Lontoc CAA, Octura JER, Iranzo MFM, Capitan SS (2014). Effects of β–Benzenehexachloride In–Ovo on the embryonic growth and development of Mallard ducks (Anas platyrhynchos domesticus L). Vet. Med. Anim. Sci. (ISSN 2054–3425).
Vrbanac BG, Stojevic Z, Milinkovic S, Balenovic T, Pirsljin J, Zdelar–Tuc M (2002). In vitro susceptibility of duck, chicken, and pig erythrocyte lipids to peroxidation. Vet. Med–Czech. 47 (10–11): 303 – 308.
Wade MG, Foster WG, Younglai EV, Mahon AC, Leingartner K, Yagminas AL, Blakey D, Fournier M, Desaulnier D, Hughes CL (2002). Effects of subchronic exposure to a complex mixture of persistent contaminants in male rats : Systemic, immune, and reproductive effects. Toxicol. Sci. 67: 131 – 143.
http://dx.doi.org/10.1093/toxsci/67.1.131
PMid:11961226
WHO (2006). WHO Food Additives Series 55, Safety Evaluation of Certain Contaminant in Food. International Programme on Chemical Safety, Geneva.
William TD, Challenger WO, Christians JK, Evanson M, Love O, Vezina F (2004). What causes the decrease in haematocrit during egg production. BES Funct. Ecol. 18: 330 – 336.
http://dx.doi.org/10.1111/j.0269-8463.2004.00829.x



