Advances in Animal and Veterinary Sciences
Research Article
Advances in Animal and Veterinary Sciences. 2 (1): 20 – 25Characterization of the Antibacterial Activity of Bacteria Associated with Stylissa Sp, a Marine Sponge
Pandian Krishnan, Malathi Balasubramaniam, S. Dam Roy, Kamal Sarma, Rizwana Hairun, Jai Sunder*
*Corresponding author: jaisunder@rediffmail.com
ARTICLE CITATION: Krishnan P, Balasubramaniam M, Roy SD, Sarma K, Hairun R, Sunder J (2014). Characterization of the antibacterial activity of bacteria associated with stylissa sp, a marine sponge. Adv. Anim. Vet. Sci. 2 (1): 20 – 25.
Received: 2013–09–28, Revised: 2013–11–13, Accepted: 2013–11–14
The electronic version of this article is the complete one and can be found online at (http://dx.doi.org/10.14737/journal.aavs/2014.2.1.20.25) which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited
Abstract
Among marine invertebrates, sponges are the most prolific phylum, with regard to presence of novel pharmacologically active compounds. Bacteria, living on the surface of marine invertebrates, have been found to produce chemicals that have potential antimicrobial activities. To test this hypothesis, a total of about 30 bacterial isolates associated with the marine sponge, Stylissa sp collected from North Bay, Andaman were successfully screened for antimicrobial activity against selected human, animal and plant pathogens including Aeromonas hydrophila, Bacillus subtilis, Enterococcus durans, Streptococcus lentus, Klebsiella pneumoniae and Rolstonia solanacearum using disc diffusion assay. The bacterial isolates were identified as belonging to the genera Marinomonas, Alteromonas, Marinobacter, Chromohalobacter, Halomonas, Cytophagas and Amylocolate based on their biochemical properties. Of the total bacterial isolates tested, 66.67% were found to have antimicrobial properties ranging from broad spectrum to species specific. One of the isolates, S8 inhibited K. pneumoniae with about 95% and 30% of the efficiency of Erythromycin and Ciprofloxacin, respectively. The findings of the study highlight the importance of sponge surface–associated bacteria in producing new natural compounds with antimicrobial activities.
Introduction
Sponges (Phylum: Porifera) are the most primitive multicellular invertebrates representing the phylogenetically oldest metazoans that evolved 750–570 million years ago (Müller 1998). Sponges have been considered as a gold mine for the chemist. More novel bioactive metabolites are obtained from sponges each year than from any other marine taxon, and a range of pharmacological properties has been demonstrated (Blunt et al. 2006). The origin of bioactive compounds in marine sponges may be a copperative interaction or a symbiosis between sponges and associated microorganisms (Flowers et al., 1998; Piel, 2004). Interestingly, in at least some cases, the compounds appear to be produced by associated microorganisms rather than by the sponge itself (Piel et al., 2004). In spite of the potential of sponge as a reservoir of many drugs, till date, not many find their way to the pharmacy shelves as approved drugs; the major issue in progress being the availability of sponges in quantities amenable for industrial extraction (Thoms and Schupp 2005). Biologically active natural products are often produced in relatively small amounts, and often by rare animals whose natural populations cannot sustain the extensive collections required for clinical trials. Thus an alternative strategy targeting the microorganisms associated with sponges for the screening of bioactive natural products may prove to be an effective approach to circumvent the associated difficulties of dealing with the organism itself. Hence, the current interest has been on the microbes associated with the sponges which can be a potential source for the bioactive compounds. Of late, marine sponges have gained importance as a rich source of compounds, possessing novel structures and exhibiting interesting biological activities (Schröder et al., 2003). Many sponge or sponge symbiont–derived metabolites are potent antibacterial, antifungal, antifeeding and antifouling compounds (Becerro et al., 1997). Some marine bioproducts originally isolated from macroorganisms, such as sponges, have been subsequently discovered to be present in microbial associates (Bewley et al., 1996). The role of sponge–associated microorganisms in the synthesis of compounds of biological interest is a subject of scientific debate, as in some cases these bacteria are reported to produce bioactive metabolites (Imhoff and Stöhr 2003). Antimicrobial activity has been reported for the bacteria associated with marine sponges collected from diverse areas viz., Antarctic region (Mangano et al., 2009), temperate (Chellossi et al., 2004) and tropical coastal waters (Thakur and Anil 2000, Kanagasabapathy et al., 2005, Anand et al., 2006). The present investigation aims at characterizing the cultivable heterotrophic fraction of surface–associated bacteria in a marine sponge, Stylissa sp from Andaman, India and investigating their antimicrobial properties against selected human, animal and plant pathogens.
MATERIALS AND METHODS
Sampling Site and Collection Of Sponges
The marine sponge, Stylissa sp (Figure 1) was collected using Self–Contained Underwater Breathing Apparatus (SCUBA) from North Bay, Andaman at a depth of 5–10 mts in December 2008. The sponge samples (250g) meant for the preparation of sponge extracts were rinsed with sterile seawater to remove adhering debris and associated biota and immediately placed in 1 lit of Methanol. For isolating sponge associated bacteria, three sponge samples were placed separately in sample boxes containing pre–sterilized seawater, cooled in an insulation box, transported immediately to the laboratory and processed within 3 hrs. Ambient seawater was also collected in sterile glass bottles and brought to the laboratory in cold chain.Fig1

Preparation of Sponge Extract
The sponge sample was allowed to soak in methanol for 2 days with intermittent shaking after which the extract was decanted and collected. The extraction process was repeated 3 times, each time by adding equal volume of fresh methanol. These extracts were pooled, filtered and concentrated under reduced pressure using a rotary evaporator. The crude sponge extract was then scrapped out and aliquots were made in phosphate buffered saline (PBS) at a concentration of 5 mg/ml. The sponge extract thus obtained was stored at 4°C till it was subjected to antibacterial activity tests.
Isolation and Identification of Sponge Associated Bacteria
Once in the laboratory, the specimens were placed separately in beakers with sterile seawater, which were changed 4 to 5 times. One gram of sponge tissue was collected using a sterile knife, and placed in a tube containing 10 ml of physiological saline (0.85% NaCl). After vigorous mixing, the samples were serially diluted up to 10–3 and 100 µl of each dilution was plated on Zobell marine agar plates. After incubating at 30°C for 48 hrs, the isolated colonies showing distinct morphology on agar plates were sub–cultured on a fresh agar plate. The isolated colonies were then streaked in slants in screw cap tubes and stored as pure cultures. A total of 62 isolates were obtained from the sponge surface, designated serially as S1–62 and stored for further screening.
These bacterial isolates were identified to the genus level using Bergey’s Manual of Determinative Bacteriology (Buchanan and Gibbons 1984). The isolates from the seawater samples were processed in a similar way and the biochemical test results were compared with that of the sponge associated bacterial isolates. Isolates from sponge samples which yielded same results as those from water samples were considered as “generalists” and were excluded from the subsequent steps for testing the antimicrobial properties.
The biochemical test results were scored as 1 if positive or 0 if negative. The hierarchical cluster analysis was done using NTSYS 2.02 software. Clustering was performed by the unweighted pair–group method of analysis (UPGMA) with statistical support.
Collection of Bacterial Metabolites
A loop full of the pure cultures of the surface associated bacteria was inoculated in nutrient broth with 1.5% salt and incubated overnight in an orbital shaker. The broth was centrifuged (10,000 rpm for 10 min) when the bacterial growth reached the exponential phase at room temperature (30oC). The supernatant was collected, concentrated 10 folds in a vacuum drier, filter sterilized using a 45µ membrane filter and the bacterial metabolite thus prepared was stored at 4oC for subsequent screening of their antimicrobial properties.
Studying the Antimicrobial Property
The sponge extracts and the bacterial metabolites were evaluated for their anatagonistic property against selected human, animal and plant pathogens viz., Aeromonas hydrophila, Enterococcus durans, Streptococcus lentus, Bacillus subtilis, Bacillus licheniformis, Streptococcus thermophilus, Klebsiella pneumoniae and Ralstonia solanacearum using disc diffusion assay (Hellio et al., 2001). A volume of 25 µl of the sponge extract and the bacterial metabolite from the sponge–associated bacterial isolates were used to saturate a sterile filter paper disc (Whatman, 6mm) and allowed to dry at room temperature. These discs were placed on the surface of the marine agar plates spread with 100 µl of the liquid cultures of each of the selected pathogens. The zone of inhibition (mm) was calculated from the edge of the disc after incubating the plates at 30oC for 24 h using standard antibiotic scale (Hi–Media, India). Discs soaked with same volume of methanol and sterile broth were used as negative controls, while those impregnated with antibiotics (Erythromycin, Ciprofloxacin; Hi–Media, India) were used as positive control. All the tests were performed in triplicate and the average zone of inhibition was calculated for each replicate assay of discs with sponge extracts / concentrated bacterial supernatants.

To determine the cumulative efficiency of a particular extract against all the pathogens, a weightage was allocated to various pathogens based on their pathogenic nature within the respective class of organisms (Table 1), which was multiplied with the minimum zone of inhibition (in mm) against the respective bacterial and sponge extract. The weighted efficiency of the particular extract against each of the selected pathogens was summed up to get the cumulative efficiency.
RESULTS AND DISCUSSION
In the present investigation, the total heterotrophic bacteria associated with the sponge, Stylissa sp was found to be 6.5 x 106 cfu gm–2. From this, a total of 62 bacterial strains which belonged to three different morphotypes were isolated. The majority were white circular (70%) followed by translucent circular (18%) and white irregular (12%) colonies. The bacterial community associated with marine sponges consists of specialist bacteria specific to a certain sponge species, sponge associated bacteria that are found on several sponge species but not in seawater and generalist marine bacteria found both on sponges and in seawater (Taylor et al., 2007). The bacteria which were specifically associated with the sponge were obtained by excluding the isolates with similar characteristics and also those present in surrounding water samples. 30 such sponge–specific isolates, out of the total 62 isolates were subsequently screened for their antimicrobial properties against selected human, animal and plant pathogens.
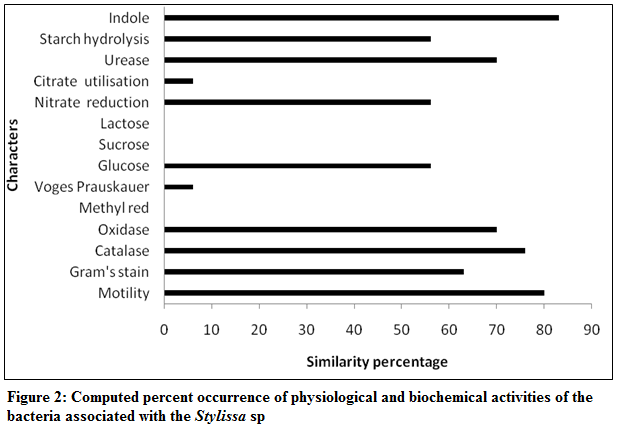
Figure 2: Computed percent occurrence of physiological and biochemical activities of the bacteria associated with the Stylissa sp
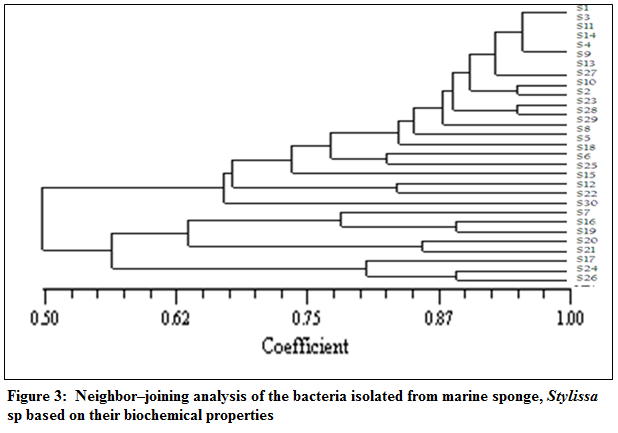
Figure 3:Neighbor–joining analysis of the bacteria isolated from marine sponge, Stylissa sp based on their biochemical properties
The bacterial isolates associated with the marine sponge, Stylissa sp were categorized based on their biochemical properties and the graphical representation of the similarity percentage is depicted in Figure 2. Over 60% of the isolates were gram positive and about 80% were motile. The results of hierarchical cluster analysis based on the biochemical tests shows that at the similarity index of 75%, the isolates could be grouped into 7 groups (Figure 3). A majority of the isolates (43%) belonged to group 1 in which 70% were found to produce antibacterial compounds. The antibacterial activity of the sponge extract was measured by disc diffusion assay which revealed that it was active against all the selected pathogens except B. subtilis. Of the selected sponge surface associated bacterial isolates, only 20 (66.67%) were found to have the antibacterial activity or in other words produce antibiotics. The antimicrobial activity of the sponge–associated bacterial isolates was more pronounced against gram positive bacteria (45%) than gram negative bacteria (15%). There was significant (P<0.05) difference in the nature of inhibitory activity of the antibacterial compounds produced by different bacterial isolates. 38% of the isolates were active against >5 pathogens, while 19% of them were active against 2–4 selected pathogens and 43% were specific to a particular pathogen. Nine bacterial isolates showed significant inhibitory activity against B. subtilis while none of the isolates was effective against R. solanacearum, a plant pathogen. Though very marginal, only isolate S6 was having antagonistic property against A. hydrophila. Among the microbes against which the antagonistic properties of the selected sponge associates were screened, S. thermophilus was inhibited by only isolates, S25–30. In the current study, maximum zone of inhibition was recorded for S8 against E. durans. The results of the antibacterial activity of the sponge isolates and their cumulative weighted efficiency are depicted in Table 2.
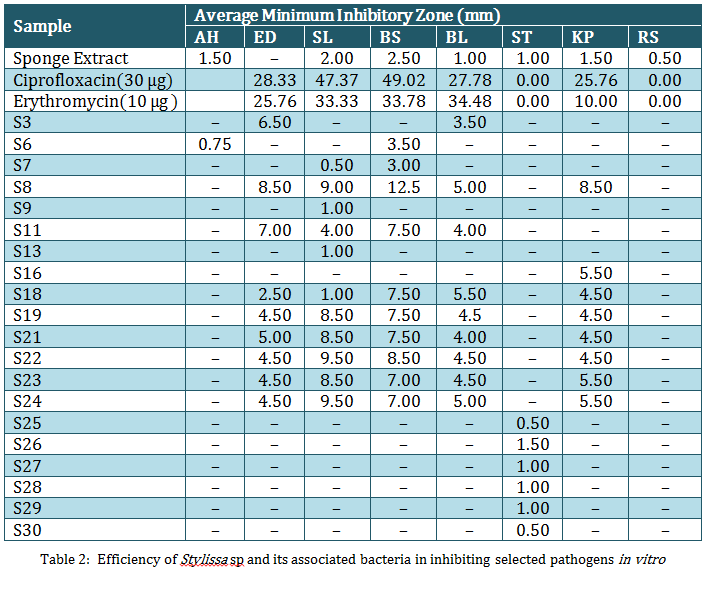
Table 2:Efficiency of Stylissa sp and its associated bacteria in inhibiting selected pathogens in vitro
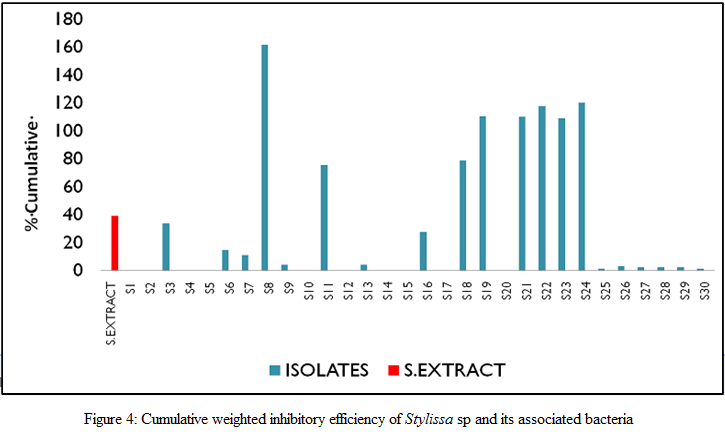
For each of the sponge associated bacteria, cumulative weighted inhibitory activity was calculated which in turn was compared against that of the sponge extract in order to determine the overall antimicrobial property of the specific isolates against selected pathogens. 26.67% of the isolates from Stylissa sp were found to have significantly (P<0.05) higher cumulative weighted inhibitory activity than the sponge extract itself (Figure 4). When expressed as the percentage of the inhibitory activity of selected antibiotics (Hi–Media discs with Ciproflaxin and Erythromycin), it was found that the isolate S8 derived from Stylissa sp had the highest percentage inhibition against all the selected pathogens. The isolate could inhibit K. pneumoniae with about 95% of the efficiency of erythromycin and 30% of that of ciproflaxin (Figure 5).
The marine environment is an exceptional reservoir of bioactive natural products, many of which exhibit structural/chemical features not found in terrestrial natural products. Marine sponges are a rich source of novel and biolgically active metabolites and are known to be the most prolific marine producers of novel compounds, with more than 200 new metabolites reported each year (Taylor et al., 2007). Andaman and Nicobar group of Islands in India constitute about 25% of the country’s coast line and are bestowed with rich marine biodiversity. Sponges are one of the biologically and ecologically significant resources of the Islands which are hitherto unexplored for their pharmaceutical potential. The islands are home for about 61 of the total 486 sponge species identified in India (Thomas 1998). The reports on antimicrobial properties of sponges from the Indian coast are very scanty (Thakur and Anil 2000, Anand et al., 2006, Gandhimathi et al., 2008). In the current study, samples of the marine sponge, Stylissa sp collected from North Bay, South Andaman were studied for their antagonistic property against selected pathogens highlighting the role of associated bacteria. The interactions between sponges and bacteria in the marine environment are poorly understood. It is generally believed that symbiotic interactions exist between sponges and microorganisms. Sponges harbor diverse and complex assemblages of microorganisms, including cultivable bacteria from seven phyla (Michael et al., 2007). Sponge host a large community of bacteria with 26 bacterial phyla discovered (Lee et al., 2011; Flemer et al., 2012). It has also been suggested that some of these bacteria chemically defend the host against microbial infection (Engel et al., 2002). Therefore, the sponge–microbes association is a potential chemical, ecological phenomenon which could provide novel pharmaceutical leads (Gandhimathi et al., 2008). Most researchers still focus on cultivable sponge associated bacterial strains that produce secondary metabolites as cultivation studies are indispensable in the discovery of novel metabolic pathways in new microbes (Michael et al., 2007). The methanol extract of Stylissa sp had the highest inhibitory efficiency against B. subtilis. This could be attributed to the cumulative inhibitory efficiency of associated bacteria, as about 35% of the sponge isolates had significant (P<0.05) antagonistic property against it. Gram positive bacteria generally predominate in the marine environment (Bernen et al., 1997), while in some recent studies gram positive and negative bacteria have been found in almost equal proportion (Anand et al., 2006). Gram positive bacteria constitute only 10% of the total isolates occurring in the Great Barrier reef (Burja et al., 1999). However, in the present study, 60% of bacterial isolates associated with Stylissa sp were found to be gram positive. The bacterial isolates from Stylissa sp belonged to the genera Marinomonas, Alteromonas, Marinobacter, Chromohalobacter, Halomonas, Cytophagas and Amylocolate. The bacterial diversity associated with sponges from South China Sea has been extensively reported by Li et al. (2007). Vibrios (West and Colwell 1984) and Pseudomonas (Thakur et al., 2003) are reported from the pelagic environment and from the surface of marine invertebrates inhabiting coastal waters. Kobayashi et al., (1994) characterized an antibiotic purified from a Vibrio sp isolated from a marine sponge, Hyrtios altum. Radhika et al., (2007) isolated Cytophagus sp, Flexibacter sp, Pseudomonas sp and Flavobacterium sp from biofilms on the databuoys installed along the Indian coast. While all of the bacterial genera reported in the Stylissa sp in the current investigation have been found to be associated with different organisms of the marine environment, their antimicrobial properties have not yet been documented. In the present study, as the antagonistic property was tested against animal, plant and human pathogens, they were given weightage ranging from 1–5 within the group. The product of weightage and the corresponding zone of inhibition against the selected pathogen yielded the cumulative inhibitory efficiency of each bacterial isolate. The cumulative inhibitory efficiency thus arrived for eight isolates (S8, S11, S18, S19, S21–24) was significantly higher than that of the sponge extract, which could be attributed to the fact that the bacterial metabolites were obtained after enriching them in growth medium while the sponge extract simply had those isolates in densities as in nature. Among them, S8 (Cytophagus sp) was found to be the most promising in terms of its antagonistic potential against the selected pathogens. Due to the increased incidence of multiple resistances among pathogenic microorganisms to drugs that are currently in clinical use, discovery of new classes of antibiotics is necessary (Burgess et al., 1999, Pompe et al., 2000). The marine sponges being the sink of many such bioactive compounds offer great scope for exploration. The present investigation identified bacterial isolates associated with Stylissa sp, which contribute to the antagonistic property of the sponge and this report being the first, to the best of our knowledge, from this geographically isolated and unique region underlines the inherent potential of exploring the marine sponges in Andamans for their bioactive properties
REFERENCES
Anand TP, Bhat AW, Shouche YS, Roy U, Siddhath J and Sarma SP (2006). Antimicrobial activity of marine bacteria associated with sponges from the waters off the coast of South East India. Micobiol. Res.161: 252–262 http://dx.doi.org/10.1016/j.micres.2005.09.002 PMid:16765842
Becerro MA, Uriz MJ and Turon X (1997). Chemically mediated interactions in benthic organisms: the chemical ecology of Crambe crambe (Porifera Poecilosclerida). Hydrobiol. 355:77–89 http://dx.doi.org/10.1023/A:1003019221354
Bernen VS, Greenstein M and Maiese WM (1997). Marine Microorganisms as a Source of New Natural Products. Advan. Appl. Microbiol. 43: 57–90 http://dx.doi.org/10.1016/S0065-2164(08)70223-5
Bewley CA, Holland ND and Faulkner DJ (1996). Two classes of metabolites from Theonella swinhoei are localized in distinct populations of bacterial symbionts. Experientia. 52: 716–722 http://dx.doi.org/10.1007/BF01925581 PMid:8698116
Blunt JW, Copp BR, Munro MH, Northcote PT and Prinsep MR (2006). Marine natural products. Nat. Prod. Rep. 23: 26–78 http://dx.doi.org/10.1039/b502792f PMid:16453031
Buchanan RE and Gibbons NE (1984). Bergey's Manual of Determinative Bacteriology. Science Publisher, 8th ed., Beijing
Burgess JG, Jordan EM, Bregu M, Mearns–Spragg A and Boyd KG (1999). Microbial antagonism: a neglected avenue of natural products research. J. Biotechnol. 70:27–32 http://dx.doi.org/10.1016/S0168-1656(99)00054-1
Burgess JG, Jordan EM, Bregu M, Mearns–Spragg A and Boyd KG (1999). Microbial antagonism: a neglected avenue of natural products research. J. Biotechnol. 70:27–32 http://dx.doi.org/10.1016/S0168-1656(99)00054-1
Burja AM, Webster NS, Murphy PT and Hill RT (1999). Microbial symbionts of Great Barrier Reef sponges. Mem Queensland Museum 44: 63–67
Engel S, Jensen PR and Fenical W (2002). Chemical ecology of marine microbial defence. J. Chem. Ecol. 28 (10):1971–1985 http://dx.doi.org/10.1023/A:1020793726898 PMid:12474894
Flemer B, Kennedy J, Margassery LM, Morrissey JP, O'Gara F, Dobson ADW (2012). Diversity and antimicrobial activities of microbes from two Irish marine sponges, Suberites carnosus and Leucosolenia sp. J. Appl. Microbiol., 112:289–301 http://dx.doi.org/10.1111/j.1365-2672.2011.05211.x PMid:22129274
Flowers AE, Garson MJ, Webb RI, Dumdei EJ, Charan RD. 1998. Cellular origin of chlorinated diketopiperazines in the dictyoceratid sponge, Dysidea herbacea (Keller). Cell Tissue. Res., 292: 597–607 PMid:9582417
Ganghimathi R, Arunkumar M, Selvan J, Thangavelu T, Sivaramakrishnan S, Kiran GS, Shanmugapriya S and Natarajaseenivasan K (2008). Antimicrobial potential of sponge associated marine actinomycetes. J. Micol. Med. 18: 16–22 http://dx.doi.org/10.1016/j.mycmed.2007.11.001
Hellio C, De La Broise D, Dufosse L, Le Gal Y and Bourgougnon N (2001). Inhibition of marine bacteria by extracts of macroalgae: potential use for environmentally friendly antifouling paints. Mar. Environ. Res. 52: 231–247 http://dx.doi.org/10.1016/S0141-1136(01)00092-7
Hentschel U, Hopke J, Horn M, Friedrich AB, Wagner M and Moore BS (2002). Molecular evidence for the uniform microbial community in sponges from different oceans. Appl. Environ. Microbiol. 68 (9):4431–4440 http://dx.doi.org/10.1128/AEM.68.9.4431-4440.2002 PMid:12200297 PMCid:PMC124103
Imhoff JF and Stöhr R (2003). Sponge–associated bacteria: general overview and special aspects of bacteria associated with Halichondria panicea, pp. 35–56. In W.E.G. Müller (ed.), Marine Molecular Biotechnology Springer–Verlag, Berlin PMid:15825639
Kanagasabapathy M, Sasaki H, Nakajima K, Nagata K and Nagata S (2005). Inhibitory activities of surface associated bacteria isolated from the marine sponge Pseudoceratina purpurea. Microbes. Environ. 20: 178–185 http://dx.doi.org/10.1264/jsme2.20.178
Kobayashi M, Aoki S, Gato K, Matsunami K, Kurosu M and Kitagawa I (1994). Marine natural products. XXXIV. Trisindoline, a new antibiotic indole trimer, produced by a bacterium of Vibrio sp. separated from marine sponge Hyrtios altum. Chem. Pharm. Bull. (Tokyo) 42 : 2449–2451 http://dx.doi.org/10.1248/cpb.42.2449
Li Z, Hu Y, Huang Y and Huang Y (2007). Isolation and phylogenetic analysis of the biologically active bacteria associated with three South China Sea sponges. Microbiol. 76: 494–499 http://dx.doi.org/10.1134/S0026261707040169
Lee OO, Wang Y, Yang J, LafiF F, Al–Sawailem A, Qian PY (2011). Pyrosequencing reveals highly diverse and speciesspecific microbial communities in sponges from the Red Sea. ISME J., 5: 650–664 http://dx.doi.org/10.1038/ismej.2010.165
Mangano S, Michaaud L, Caruso, C, Brilli M, Bruni V, Fani R and Giudice AL (2009). Antagonistic interactions between psychrotrophic cultivable bacteria isolated from Antarctic sponges: a preliminary analysis. Res. Microbiol. 160: 27–37 http://dx.doi.org/10.1016/j.resmic.2008.09.013 PMid:19013518
Michael WT, Regina R, Doris S, Michael W (2007). Spongeassociated microorganisms: evolution, ecology and biotechnological potential. Microbiol. Mol. Biol. R., 71: 295–347 http://dx.doi.org/10.1128/MMBR.00040-06 PMid:17554047 PMCid:PMC1899876
Müller WEG (1998). Origin of Metazoa: Sponges as living fossils. Naturwissenschaften. 85: 11–25 http://dx.doi.org/10.1007/s001140050444
Piel J, Hui D, Wen G, Butzke D, Platzer M, Fusetani N and Matsunaga S (2004). Antitumor polyketide biosynthesis by an uncultivated bacterial symbiont of the marine sponge Theonella swinhoei. Proc. Natl. Acad. Sci. USA 101: 16222–16227 http://dx.doi.org/10.1073/pnas.0405976101 PMid:15520376 PMCid:PMC528957
Piel J (2004). Metabolites from symbiotic bacteria. Nat. Prod. Rep., 21: 519–538 http://dx.doi.org/10.1039/b310175b PMid:15282634
Pompe S, Simon J, Wiedemann PM and Tannert C (2005). Future trends and challenges in pathogenomics. EMBO. Rep.: 6, 7: 600–605 http://dx.doi.org/10.1038/sj.embor.7400472 PMid:15995675 PMCid:PMC1369123
Radhika G, Venkatesan R and Kathiroli, S (2007). N–methylpyrrolidone: Isolation and characterization of the compound from the marine sponge Clathria frondifera (class: Demospongiae). Ind. J. Marine Sci. 36: 235–238
Schröder HC, Brümmer F, Fattorusso E, Aiello A, Menna M, de Rosa S, Batel R and Müller WEG (2003). Sustainable production of bioactive compounds from sponges: primorphs as bioreactors. In: Müller WEG (ed) Marine Molecular Biotechnology. Springer–Verlag, Berlin, p 163–197
Taylor MW, Radax R, Steger D and Wagner M (2007). Sponge–Associated Microorganisms: Evolution, Ecology and Biotechnological Potential, Microbiology and molecular biology reviews. 71(2): 295–347 http://dx.doi.org/10.1128/MMBR.00040-06 PMid:17554047 PMCid:PMC1899876
Thakur NL and Anil AC (2000). Antibacterial activity of the sponge Ircinia ramosa: importance of its surface associated bacteria. J. Chem. Ecol. 26: 57–71 http://dx.doi.org/10.1023/A:1005485310488
Thakur NL, Anil AC and Müller WEG (2004). Culturable epibacteria of the marine sponge Ircinia fusca: temporal variations and their possible role in the epibacterial defense of the host. Aquatic. Microb. Ecol. 37: 295–304 http://dx.doi.org/10.3354/ame037295
Thakur NL, Hentschel U, Krasko A, Pabel CT, Anil AC and Müller WEG (2003). Antibacterial activity of the sponge Suberites domuncula and its primorphs: potential basis for epibacterial chemical defense. Aquat. Microb. Ecol. 31: 77–83 http://dx.doi.org/10.3354/ame031077
Thomas PA (1998). Faunal diversity in India. A commemorative volume in the 50th years of India's independence. In: Alfred JRB, Das AK and Sanyal AK, Editors, 1998. Envis Center, Zoological Survey of India, Calcutta, pp. 28–36
West PA and Colwell RR (1984). Identification and classification of Vibrionaceae – an overview. In: Colwell, R.R. (Ed.), Vibrios in the Environment. Wiley, New York, pp. 285–363



