Journal of Infection and Molecular Biology
Review Article
Journal of Infection and Molecular Biology 2 (4): 61 – 73Pseudomonas Aeruginosa Biofilm: Role of Exopolysaccharides in the Function and Architecture of the Biofilm
Aamir Ghafoor1*, Aamir Ghafoor1*, Zain–ul–Fatima2, Aqeel Javed3
- University Diagnostic Lab, University of Veterinary and animal Sciences, Lahore Pakistan
- Department of Pathology, University of Veterinary and animal Sciences, Lahore Pakistan
- Department of Pharmacology, University of Veterinary and animal Sciences, Lahore Pakistan
*Corresponding author:aamir.ghafoor@uvas.edu.pk
ARTICLE CITATION:
Ghafoor A, Zain–ul–Fatima, Javed A (2014). Pseudomonas aeruginosa biofilm: role of exopolysaccharides in the function and architecture of the biofilm. J. Inf. Mol. Biol. 2 (4): 61 – 73.
Received: 2014–04–28, Revised: 2014–09–08, Accepted: 2014–09–15
The electronic version of this article is the complete one and can be found online at
(
http://dx.doi.org/10.14737/jimb.2307–5465/2.4.61.73
)
which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited
ABSTRACT
More than 90% bacteria live as a biofilm in natural habitats. This surface–attached, sessile mode of life confers many advantages to bacteria and increases their survival. Pseudomonas aeruginosa, an opportunistic pathogen, has been extensively studied as a model organism for biofilm studies. Biofilm formation is a stepwise process in which motile bacteria attach to a surface and initiate a cascade of changes. Consequently these bacteria become phenotypically different from their planktonic counterparts. Initially these bacteria attach reversibly to the surface via hydrophobic interactions between the bacterial surface proteins and the substratum. In response to environmental cues, intracellular concentration of a secondary messenger, c–di–GMP, increases in the bacterial cells. This increase in c–di–GMP results in loss of motility, expression of specific adhesins, and irreversible attachment of the bacteria. These bacteria start proliferating while residing in close proximity. The bacterial cells sense the presence of other community members in their close proximity, and start communicating with each other through a mechanism known as quorum sensing. These bacterial cells produce and recruit extracellular polymeric substances (EPS) which encase the bacteria and protect them from unfavourable environmental conditions, predators and biocidal compounds. Components of the EPS include eDNA, proteins, lipids and exopolysaccharides. The exopolysaccharides produced and secreted by the bacteria play a vital role in biofilm formation. Pseudomonas aeruginosa produces various types of exopolysaccharides, but three of them are considered very important in biofilm formation; mannose–rich Psl, glucose–rich Pel, and uronic acid–rich alginate. These three exopolysaccharides are synthesized and secreted by the protein–complex encoded by their respective operons. The role played by these exopolysaccharides in architecture and function of biofilm is currently under extensive research. This review presents the mechanism of biofilm formation and the role of exopolysaccharides in biofilm.
INTRODUCTION
For a long time in the history of microbiology, microorganisms were considered as freely suspended cells in nutritionally rich media and were characterized on the basis of their growth characteristics in the media provided. Heukelekian and Heller (1940) showed for the first time that the presence of a surface for attachment substantially enhanced the bacterial growth. A subsequent study demonstrated that the number of bacteria on the surface were more as compared to their surrounding environment. These indirect evidences provided clues that bacteria can grow as a community after attaching to the surface (Zobell, 1943). This community of surface–attached bacteria is now known as biofilm. However, a detailed study of biofilm was made possible only after the use of electron microscopy. (Jones et al., 1969) used electron microscopy to show that biofilm formed in seawater on a trickling filter was composed of many different kinds of organisms. With the help of polysaccharide–specific stains, these workers also provided evidence for the presence of polysaccharides in extracellular matrix of bacterial biofilms. In subsequent years it was shown that microbial biofilms in industrial waste water were not only very persistent but also highly resistant to disinfectants such as chlorine (Characklis 1973). Later, a theory was proposed which elucidated the mechanism through which microorganisms attach to a surface and the advantages amassed by the microorganisms from this ecologic niche (Costerton et al., 1978). In the last two decades, the use of scanning electron microscopy (SEM), confocal laser scanning microscopy (CLSM) and molecular biology techniques to study gene regulation have helped to increase our understanding of architecture and functioning of biofilms.
MICROBIAL BIOFILMS
Biofilm is a consortium of microbes which is irreversibly attached (not removed by gentle washing) to a surface and enclosed in a matrix of extracellular polymeric substances (EPS). The EPS constitute 50– 90% of the total organic mass of the biofilm and is composed of exopolysaccharides, extracellular DNA (eDNA), proteins, lipids and humic substances (Flemming H–C et al., 2000). The extracellular polymeric substances encased microbescells form microcolonies which are separated from each other by the presence of water channels (Lewandowski, 2000). These water channels allow liquid to flow through them which facilitates transfer of nutrients, oxygen, waste metabolites and even antimicrobial agents into the surrounding microcolonies.
It is suggested that more than 90% of bacteria live in biofilm mode (Costerton et al., 1995; Geesey et al., 1977). Inside the biofilm, cells living in close proximity can have an increased rate of gene transfer, well developed cooperation and quorum sensing [reviewed in (Donlan, 2002)]. Furthermore, compared to planktonic bacteria, biofilms can better endure the negative consequences of pH changes, nutrient scarcity, free radicals, biocides and antimicrobial agents [reviewed in (Costerton et al., 1995)].
Importance of Microbial Biofilms
Microbial biofilms impact human life in different ways. Bacterial biofilms on medical devices such as catheters, endoscope tubes, contact lenses and implanted prosthetic devices, are a known source of nosocomial infections [reviewed in (Starkey et al., 2009)]. Microorganisms form biofilms on internal and external human surfaces and cause various infections including dental plaques, skin infections, urinary tract infections and chronic lung infections [reviewed in (Costerton et al., 1999). Recent studies have shown that chronic lung infections in cystic fibrosis (CF) patients and some persistent wound infections are due to a biofilm forming microorganism, Pseudomonas aeruginosa (Bjarnsholt et al., 2009; DeVault et al., 1990; Harmsen et al., 2010; Kirov et al., 2007).
Biofilm forming microbes also cause infections in animals. Otitis (both, media and externa), a chronic and 'hard to eradicate' infection, is caused by P. aeruginosaand Staphylococcus intermedius in dogs (Colombini et al., 2000; Petersen et al., 2002). The persistent nature of chronic mastitis in dairy animals is attributed to the ability of the microorganisms to form biofilms [reviewed in (Melchior et al., 2006)]. Biofilms in the food processing industry have also been recognized as a big problem. Bacteria can adhere and grow as biofilm in pipelines. These biofilms are difficult to eradicate due to their resistant nature, and become a continuous source of bacterial contamination [reviewed in (Poulsen, 1999)].
Microbes also form biofilms in the gastrointestinal tract of humans and animals. These biofilms provide an ecological niche where bacteria can interact with each other and with the host cells. These biofilms develop a relationship with the host which may be beneficial or harmful for the host [reviewed in (Macfarlane and Dillon, 2007)] .
These diverse effects of biofilms on human and animal lives described above have attracted a lot of attention by researchers. Although many different bacterial species have been used to study the architecture, function, regulation and control of biofilms, a lot of research has been conducted using P. aeruginosaas a model organism. Medical relevance, ubiquity, and ability to survive in diverse environments have made P. aeruginosaan ideal organism for biofilm study. In this review, salient features of biofilm formation are discussed, focusing on P. aeruginosaas the model organism.
The Model Organism: Pseudomonas aeruginosa
P. aeruginosais a gram negative, rod shaped bacterium which has an ability to adapt to a wide range of environments. In the presence of water the bacterium can colonize both biotic and abiotic surfaces. P. aeruginosais an opportunistic
pathogen which can infect plants, insect, worms and mammals (Hendrickson El Fau – Plotnikova et al., 2001; Sibley et al., 2008; Tan et al., 1999). A variety of virulence factors produced by P. aeruginosahelp it to survive in wide range of hosts. In addition, P. aeruginosaincreases the chance of its survival by living both planktonic and sessile modes of life and can switch from one mode of life to the other in response to changing environmental conditions. The ability of P. aeruginosato cause acute and chronic infections in immuno–compromised patients can be linked with planktonic and sessile modes of life respectively. Both modes of life are governed by various transcriptional regulators produced by the bacterium (Stover et al., 2000). During planktonic life, motile bacteria produce numerous virulence factors which help the bacteria to cause acute infections in the host. These factors include type IV pili (West et al., 1994; Wolfgang et al., 2003), the Type III secretion system (T3SS) (Wolfgang et al., 2003), the Type II secretion system (T2SS) secreted toxins and proteases. On the other hand, P aeruginosa can also become non–motile and starts developing a biofilm which protects bacteria from host response and increases its chance of survival inside the host. This sessile mode of life is a major cause of chronic infections in human and animals. A better understanding of this bacterial mode of life will lead us to develop better treatment and control strategies.
BIOFILM FORMATION
Biofilm formation is a stepwise, complex and dynamic process which involves many physiological, chemical, genetic and biological processes. Biofilm formation can be roughly divided into the following steps: reversible and irreversible attachment to substratum surface, followed by microcolony and biofilm formation with the help of EPS (produced and recruited by constituent bacterial cells), and finally dispersal of biofilm. A schematic representation of biofilm formation in P. aeruginosais presented in Figure 1.
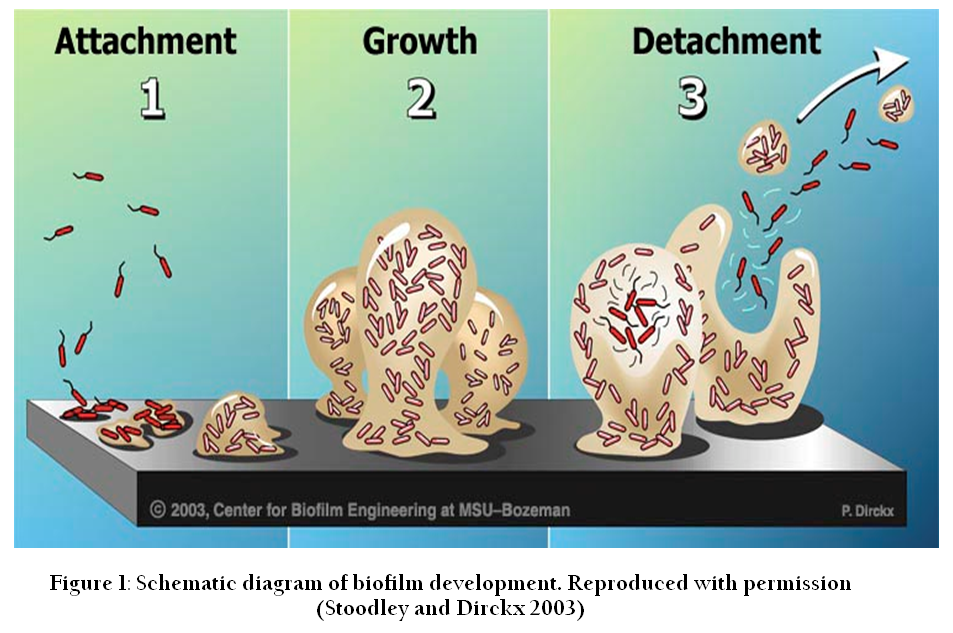
Figure 1: Figure 1: Schematic diagram of biofilm development. Reproduced with permission (Stoodley and Dirckx 2003)
Attachment to the Surface
Biofilm formation is initiated by attachment of free living cells to a surface. With the help of Brownian movement and flagellar–dependent motility, bacterial cells reach to an appropriate surface described below.
The Role of Substratum in Attachment
Compared to growth in liquid, attachment to a surface provides several advantages to the bacterial cells. First, surfaces provide a good nutritional supply because most of the macromolecules are adsorbed to the surface (called "conditioning" of the surface) [reviewed in (Donlan, 2002)]. Second, laminar flow velocity and shear forces are minimal just above the surface at the liquid–solid interface, which allows settlement of the bacterial cells on the surface [reviewed in (Donlan, 2002; Korber et al., 1989)]. Physical and chemical properties of the surface may play an important role in attachment, as rough surfaces provide an extended surface area and friction for strong attachment (Characklis, 1990). Similarly, hydrophobicity and hydrophilicity of the surfaces also play a role in bacterial attachment. Previous studies suggest that microorganisms readily attach to hydrophobic surfaces, such as Teflon and plastics, as compared to hydrophilic surfaces, such as glass and metals (Bendinger et al., 1993; Fletcher and Loeb, 1979; Pringle and Fletcher, 1983).
Environmental Cues for Attachment
Bacterial attachment to surfaces is initiated in response to some environmental cues. Scarcity of nutrition is one of the important environmental cues in response to which many bacterial species initiate attachment (Allegrucci et al., 2006; Costerton et al., 1995; Humphrey et al., 1983). Limited supply of inorganic phosphate (Pi) is sensed by the bacteria for initiation of biofilm, but a concentration of Pi below a certain threshold level impacts negatively and results in impaired attachment in Pseudomonas aureofaciens PA147–2 (Monds et al., 2001). In Escherichia coli K–12, different nutritional sources were tested and it was shown that carbon sources which were metabolized to produce acetyl coenzyme A, acetyl phosphate, and acetate resulted in initiation of biofilm formation (Prüß et al., 2010). The presence of monosaccharides (the building blocks of extracellular polysaccharide) in the growth media is sensed by Vibrio cholerae as a cue to form biofilm (Kierek and Watnick, 2003). In addition, osmolarity, divalent cations and pH are also environmental factors which are sensed as cues by many bacterial species to initiate attachment and begin biofilm formation [reviewed in (Petrova and Sauer, 2012)].
Reversible Attachment
Attachment can either be reversible or irreversible. The reversible attachment is a transitory phase and bacterial cells can detach from the surface at any time to become freely swimming cells again. Reversibly attached bacterial cells adhere to a surface via a single pole using a flagellum, fimbriae or other adhesins produced by the bacterium [reviewed in (Petrova and Sauer, 2012)]. This attachment is initiated when bacterial cells make first contact with the surface and overcome the repulsive electrostatic forces. A repulsion is expected, due to net negative charges both on bacterial and other surfaces [reviewed in (Goulter et al., 2009; Pratt and Kolter, 1999)]. However, these repulsive forces are overcome by the hydrophobic interactions between adhesins present at the bacterial surface and the attachment surface (Corpe, 1980). The role of fimbriae in bacterial attachment to surfaces has been reported by many studies (Bullitt and Makowski, 1995; Rosenberg et al., 1982) and most fimbriae have been shown to contain high numbers of hydrophobic amino acid residues and may contribute to cell surface hydrophobicity (Rosenberg et al., 1982). Flagella have also been shown required for attachment.
In P. aeruginosa, mutations in flagellar biosynthesis and type IV pili genes rendered the mutants defective in attachment to the surface (O'Toole and Kolter, 1998). It was speculated that this defect in attachment was due to inability of the mutants to swim and reach the surface. However, later studies proposed that FilD, a flagellar cap protein, interacts with the mucin and may be required for attachment in the lungs of cystic fibrosis (CF) patients (Arora et al., 1998). Recent studies have shown that attachment is reduced in the absence of structural flagellar proteins and defects in flagellar motility also results in impaired attachment and reduced biofilm growth. P. aeruginosahave a single flagellum and one motor but two stators, MotAB and MotCD, which provide energy for flagellar rotation. Although the presence of a single stator was sufficient for flagellar movement, both were shown to be essential for attachment and biofilm development. Using beads coated with purified Type IV pili, a recent study has demonstrated that type IV pili are essential and sufficient for maximal binding to the surface of polarized epithelial host cells (Bucior et al., 2012). In addition, a chaperone–usher pathway (Cup) which synthesizes and assembles the fimbriae also has been suggested as an essential component for attachment to abiotic surfaces (Ruer et al., 2007).
Irreversible Attachment
Irreversibly attached bacteria remain fixed to the surface and form a monolayer. In the case of irreversible attachment, cells are adhered to a surface along their long axes and show reduced flagellar movements [Reviewed in (Petrova and Sauer, 2012)]. A few factors have been identified in some bacterial species which are suggested to be involved in the transition from reversible to irreversible attachment state.
The transition from reversible to irreversible attachment in P aeruginosa has been linked with SadB/SadC/BifA protein complexes. These proteins control swarming motility and biofilm formation [reviewed in (Petrova and Sauer, 2012)]. In many bacteria transition from planktonic to sessile mode of life is controlled by a secondary messenger known as cyclic diguanylate monophosphate (c–di–GMP) which is synthesized from two molecules of GTP. The group of proteins which synthesize c–di–GMP are known as diguanylate cyclases (DGCs) and the group of proteins which are involved in the degradation of this secondary messenger are known as phosphodiesterases (PDEs) (Hengge, 2009). In P aeruginosa, SadC is a DGC which synthesises c–di–GMP in a localized area and, as a result of that, motility is reduced and biofilm development is increased (Merritt et al., 2007; Merritt et al., 2010). In contrast, BifA, a PDE, c–di–GMP degrading enzyme, increases motility of bacterial cells, decreases attachment and reduces biofilm development (Merritt et al., 2007; Merritt et al., 2010). SadB is suggested to control the transition from reversible to irreversible by decreasing flagellar reversal rate and increasing exopolysaccharide production (Caiazza et al., 2007; Caiazza and O'Toole, 2004).
In Pseudomonas fluorescens, a large protein, LapA, is secreted which is associated with the cell surface and helps the bacterial cell to attach permanently to surfaces (Hinsa et al., 2003).Similarly, another large protein, LapF, secreted by Pseudomonas putida is required for attachment and colonization on the roots of the plants (Martínez–Gil et al., 2010). Interestingly no orthologues of LapA and LapF are found in P. aeruginosa (Fuqua 2010) but presence of large protein, CdrA (c–di–GMP–regulated protein A), has been suggested to be required to form stabilized biofilm structures by interacting with Psl(Borlee et al., 2010). In E. coli, biosynthesis and secretion of poly–δ–glutamate (PGA) is proposed as an important adhesin required for permanent attachment (Agladze et al., 2005). In Caulobacter crescentus transition is initiated by removal of flagellum with the help of protease (Kanbe et al., 2005) and an exopolysaccharide 'holdfast' is produced which is composed of oligomers of N–acetylglucosamine (Janakiraman and Brun, 1999; Smith et al., 2003). The holdfast acts as an adhesin and firmly attaches the bacterium to the surface (Tsang et al., 2006).
Microcolony and Biofilm Formation
In some microbes, transition from reversible to irreversible attachment is mediated by the presence of EPS. After attachment, the bacterial cells start proliferating to form localized microcolonies and EPS production increases within the microcolonies. Eventually a hydrated extracellular matrix encases the immobilized bacterial cells, where bacterial cell division continues inside the matrix and results in formation of a heterogeneous biofilm [reviewed in (López et al., 2010; O'Toole et al., 2000)]. Increase in heterogeneity of biofilm allows the development of physical and chemical microgradients inside the biofilm, including those of nutrients, pH and oxygen [reviewed in (Stewart and Franklin, 2008)]. For instance, when biofilm attains a thickness of 10–25µm, the microenvironment near the attachment surface changes and becomes anaerobic (Hamilton, 1987). This decreased oxygen supply may result in altered gene expression in the cells. Many subpopulations may develop within the biofilm with differential gene expression in different subpopulations of the biofilm. For instance, a study showed that enzymatic activity of an oxygen–dependent alkaline phosphatase was high at the surface and low at the base of P. aeruginosabiofilm (Xu et al., 1998). This was related to less oxygen supply near the base compared to that on the surface. Similarly, protein and DNA synthesis was observed in different subpopulation of S. aureus biofilm. The cells located near the surface with ample supply of oxygen showed high metabolic activity, whereas the cells close to agar were metabolically less active (Rani et al., 2007). This presence of sub–populations of cells within biofilm is an indicative of division of labour. This division of labour between cells in the biofilm also influences the architecture of the biofilm (Barken et al., 2008).
The architecture of hydrated biofilm was impossible to study using light or electron microscopy, but became evident using CLSM. Many studies have demonstrated that biofilm are composed of clustered cells, encased in an extracellular matrix. On the basis of the shape of these cell clusters, biofilm architecture can be designated as flat, filamentous, cylindrical or mushroom–like [reviewed in (Flemming et al., 2007; Flemming and Wingender 2010)]. Architecture of the biofilm is also influenced by availability of nutrients. The wild–type P. aeruginosaPAO1 usually forms mushroom–like microcolonies. However, when genes required for the iron binding protein pyoverdine were inactivated, the mutants formed very thin and unstructured biofilms even in the presence of iron in the media (Banin et al., 2005). Increase in iron supply above a threshold level also proved to be harmful for microcolony formation in wild type P. aeruginosaPAO1 (Yang et al., 2007).
Extracellular Matrix Formation
EPS make a meshwork of polymers which contribute to cell to surface attachment, cell to cell attachment, and provide a scaffold for the architecture of the biofilm. The EPS hold the cells together and help them to live in close proximity where cell to cell communication is made possible; this is essential for the functioning of the biofilm. EPS protect bacterial cells from desiccation, biocidal agents, protozoal grazing, heavy metals, free radicals and the host immune system [reviewed in (Flemming and Wingender, 2010)].
The Matrix is composed of EPS, metal ions and water. Initially the extracellular polysaccharides were thought to be the only polymeric substances present in the biofilm matrix, but many other components have subsequently been identified. Components of the EPS are eDNA, proteins, lipids, polysaccharides and humic substances [reviewed in (Branda et al., 2005; Flemming and Wingender, 2010)].
Extracellular DNA
Previously, eDNA was considered as a bi–product of lysed cells, but recent studies have shown that eDNA is a very important polymer in the matrix of biofilms formed by both Gram–positive and Gram–negative bacteria (Allesen–Holm et al., 2006; Qin et al., 2007). eDNA helps to maintain the architecture of the biofilm and acts like glue to stabilize the cell to cell connections (Whitchurch et al., 2002). A role of eDNA has also been suggested in protection of bacterial cells in biofilm against the harmful effects of cationic antibacterial polypeptides (CAP) (Mulcahy et al., 2008). In bacterial biofilms, eDNA is discharged by various mechanisms including prophage–mediated cell death (Kirov et al., 2007; Webb et al., 2003), quorum sensing regulated DNA release (Allesen–Holm et al., 2006), and by outer membrane vesicles (OMV) produced by living cells (Kadurugamuwa and Beveridge, 1995; Schooling and Beveridge 2006). In P. aeruginosa, eDNA plays an important role in architecture and function of the biofilm (Whitchurch et al., 2002). Treatment of P. aeruginosabiofilms with DNase at early stages of biofilm development dissolved the biofilm. However, DNase treatment at later stages did not prove detrimental (Whitchurch et al., 2002). In a young two days–old P. aeruginosabiofilm, eDNA was located on the surface of microcolonies, whereas it was found around the stalk of mushroom–like biofilm structures in the mature biofilm (Allesen–Holm et al., 2006). Recently eDNA has been shown as important component for the motility of the P. aeruginosacells. It acts as trails and allows cells to move on it and this helps bacterial biofilms to organise their cells in the required way to develop the architecture of the biofilm (Gloag et al., 2013).
The role of eDNA has been studied in various other bacterial biofilms. In Haemophilus influenzae biofilms, eDNA forms a dense meshwork of fine threads like a 'thick rope' that span water channels (Jurcisek and Bakaletz, 2007). Members of the genus Rhodovulum form self–flocculating, aggregating structures with the help of exopolysaccharides, proteins and eDNA. Interestingly, DNase treatment of these clumps resulted in breakdown of the structures, whereas protein and polysaccharide lysing enzymes showed no impact on biofilm architecture, suggesting eDNA as the main polymer involved in biofilm formation in Rhodovulum (Watanabe et al., 1998).
Proteins
Proteins are also important structural components of the extracellular matrix in biofilm. In addition, some enzymes are also held in EPS which make an external digestive system for bacterial cells.
In P. aeruginosa, twogalactose–specific lectins, LecA and LecB, have been shown important for the maintenance of the biofilm architecture (Diggle et al., 2006; Tielker et al., 2005). Biofilm was completely disrupted when LecB was targeted with high–affinity ligand (Johansson et al., 2008). Role of another polymeric protein, CdrA, has also been demonstrated for P aeruginosa biofilm. CdrA binds with exopolysaccharide Psl, and links Psl with Psl or Psl with cell (Borlee et al., 2010). In addition, Cup fimbriae (Vallet et al., 2001) and type IV pili (Klausen et al., 2003) are also required for architecture of the P. aeruginosabiofilm. Similarly, TasA in B. subtilis (Branda et al., 2006) and Bap in S. aureus (Lasa and Penades, 2006) are two more examples of proteins which are required to form biofilms by the respective bacteria species.
Exopolysaccharides
Many bacteria produce extracellular polysaccharides that play important roles in bacterial virulence. These extracellular polysaccharides can be classified into two subclasses: (a) capsular polysaccharides, which remain associated with the cells after harvesting the bacteria by centrifugation, and (b) exopolysaccharides which remain in the supernatant. This distinction can be well defined for bacteria grown in shaken cultures, but it is difficult to discriminate between the two when studying structured communities such as biofilms. Nevertheless, many studies strongly support the notion that biofilm architecture is mainly defined by these extracellular polysaccharides produced by the biofilm–forming bacterial species [reviewed in(Branda et al., 2005)].
Cellulose is one of the most studied exopolysaccharide regarding biofilm formation. Gluconacetobacter xylinus is the first species shown to produce extracellular cellulose (Ross et al., 1987). Cellulose has also been reported in some other bacterial species, for instance Salmonella typhimurium and Escherichia coli. (Zogaj et al., 2001). Comparative genome analysis of many other species has revealed the presence of genes homologous to- the G. xylinus BCS (Bacterial Cellulose Synthesis), indicating their capability to produce cellulose (Ross et al., 1991). Cellulose is shown to be involved in biofilm formation in such bacterial species. It forms gel which retains water and consequently protects the residing bacteria from desiccation (Yamanaka et al., 1989).
In addition to cellulose, there are other exopolysaccharides known to be involved in the formation of extracellular matrices. For example, the exopolysaccharide intercellular adhesin (PIA), produced by enzymes encoded by the ica locus in staphylococcal species (Götz 2002). PIA synthesized by Staphylococcus epidermidis and S. aureus is also known as poly–N–acetyl glucosamine (PNAG) polymer. These polymers act as adhesins, and are essential for biofilm architecture (Mack et al., 1996; Mack et al., 1994; Maira–Litrán et al., 2002). PIA–like polymers were also reported to be synthesized in other bacterial species. For example in E. coli MG1655, a polymer similar to PIA has been shown to play a role in biofilm formation (Wang et al., 2004). Similarly, in Yersinia pestis, synthesizes a polymer similar to PIA which is required for the biofilm formation inside the mouth of parasite host which results in an irritation in parasite and the parasite bites to the human host. Consequently results in spreading of plague (Darby et al., 2002). Colanic acid is another sugar polymer produced by E. coli , composed of galactose, fructose and glucose,and commonly found in extracellular matrix of biofilms formed by E coli(Danese et al., 2000).
Most strains of the Gram–positive bacterium B. subtilis are capable of synthesizing two polymers: the exopolysaccharides, EPS and PGA, which play important role in the process of biofilm formation (Branda et al., 2006; Stanley and Lazazzera, 2005). The role played by each polymer depends on the strain and environment surrounding the biofilm. For instance, the wild strain NCIB3610 needs exopolysaccharide EPS1 but not PGA to form the biofilm (Branda et al., 2006). PGA overproducing mutants formed mucoid colonies and biofilm which lost its structure. In some wild strains like RO–FF–1 and the laboratory strain JH642, PGA is naturally produced and is crucial for surface–attached biofilm formation (Stanley and Lazazzera, 2005).
Teichoic acid is another exopolysaccharide which constitutes the biofilm matrix in S. epidermidis (Sadovskaya et al., 2004). S. epidermidis synthesizes two types of teichoic acid: one which is linked with the bacterial membrane, and the other which is secreted into extracellular milieu. Secretion of the extracellular teichoic acid increases viscosity of the colony [reviewed in (Lembre et al., 2012)].
P. aeruginosahas been extensively used as a model organism to study biofilms. Three exopolysaccharides important in biofilm formation in P. aeruginosaare Psl, Pel and alginate. Psl is a mannose–rich exopolysaccharide, which has been shown essential for the initial attachment and biofilm architecture (Ma et al., 2007). The Role of Pel, likely to be glucose–rich exopolysaccharide (Friedman and Kolter, 2004), is not well understood yet. Alginate is an anionic polymer of manuronic acid and gluronic acid and plays a very important role in biofilm architecture and function (Hay et al., 2009).
All above mentioned polysaccharides play key role in biofilm formation. A detailed understanding of these exopolysaccharides will help us to understand the 'building blocks' of biofilm. A detailed account of the exopolysaccharides produced by P. aeruginosaand their role in biofilm formation and function is given in the sections to follow (for Psl and Pel in chapter I, section 2, and for alginate in chapter II).
Dispersal of Biofilm
After maturation, the biofilm continues its life cycle by dispersing bacterial cells. After leaving the mature biofilm, these planktonic cells again attach to a new surface and start the development of another microcolony. Many studies have identified that the dispersal is initiated in response to various external environmental cues. Changes in availability of the following nutrients have been identified as signals that trigger dispersal: a) carbon sources,b) oxygen, c) nitric oxide (NO), and d) iron. In addition, many signalling molecules including the quorum sensing (QS) signalling molecule AHL, diffusible fatty acids, peptides and D– amino acids have been found responsible for initiation of the process [reviewed in (McDougald et al., 2012)] .
In P. aeruginosa, and other Pseudomonas species, active dispersal of the bacteria from biofilm is initiated by localized cell death in a subpopulation (Tolker–Nielsen et al., 2000; Webb et al., 2003). These dead cells are used as nutrients by other bacterial cells which provide these cells with sufficient energy to become 'dispersal cells' [reviewed in (McDougald et al., 2012)]. These active cells leave the biofilm through 'local break out points' and swim away to start another lifecycle (Purevdorj–Gage et al., 2005). In P aeruginosa, localized cell death is also thought to be a source of digestive enzymes which might play a role in the digestion of the exopolysaccharides and allow the release of bacterial cells [reviewed in (McDougald et al., 2008)].
The intracellular level of c–di–GMP controls the mode of life in bacteria. Decrease in concentration of this secondary messenger leads to changes which result in switching from sessile to planktonic life again (Barraud et al., 2009). Nitric Oxide has been identified as a signal in P. aeruginosawhich activates the PDEs. Activated PDEs can lead to a decrease in c–di–GMP level in bacterial cell. Consequently the bacterial cells become motile and ready to disperse (Barraud et al., 2009).
REGULATION OF BIOFILM FORMATION
Two important molecules involved in regulation are discussed below.
Cyclic Diguanylate Monophosphate
In the bacterial cell c–di–GMP is synthesized by DGCs from two molecules of GTP and degraded to 5′–phosphoguanylyl –(3′–5′)–guanosine (pGpG) or guanosine monophosphate (GMP) by PDEs (Hengge 2009). DGCs and PDEs are characterized by the presence of the conserved domain GGDEF (Paul et al., 2004) and EAL / HD–GYP (Christen et al., 2005) (Ryan et al., 2006), respectively. Proteins containing GGDEF/EAL domains are mostly multi–domain proteins that contain some regulatory domains (Galperin et al., 2001). These regulatory domains respond to environmental stimuli and consequently induce the DGC or PDE activity of the protein. In response to increased activity of DGCs and PDEs, localized concentration of c–di–GMP is changed. The effector proteins, containing specific c–di–GMP– binding domains, bind to c–di–GMP (Ryan and Dow, 2010). The effectors become activated or inactivated in response to binding with c–di–GMP and regulate motility, attachment, exopolysaccharide biosynthesis, biofilm formation and dispersal of the biofilm [reviewed in (Hengge, 2009)].
In P. aeruginosa, c–di–GMP plays its regulatory role in all steps of biofilm development, from attachment to dispersal. Flagellar and Type IV pili, important structures for initial attachment, are regulated by GGDEF/EAL–containing proteins such as MorA and FimX. MorA mutants showed an inability to form biofilm whereas FimX mutants lost polarity of pili expression (Choy et al., 2004; Huang et al., 2003). As mentioned above (Page No. 8), SadC increases c–di–GMP, thisresults in the transition from reversible to irreversible attachment (Merritt et al., 2007; Merritt et al., 2010). C–di–GMP controls the biosynthesis of exopolysaccharide at transcriptional and translational levels. FleQ represses the transcription of pel operon after binding to its promoter but, in the presence of c–di–GMP, FleQ is inactivated and the promoter is derepressed (Baraquet et al., 2012). Pel biosynthesis is controlled at post–transcriptional level by c–di–GMP–binding protein, PelD (Lee et al., 2007). C–di–GMP also regulates the exopolysaccharide alginate biosynthesis in P. aeruginosa(Hay et al., 2009). An important protein involved in alginate biosynthesis, Alg44, binds with c–di–GMP for its activity (Merighi et al., 2007).
Quorum Sensing
After irreversible attachment, the bacterial cells residing close to each other start forming microcolonies. Bacterial cells within these microcolonies communicate with each other and develop coordination amongst the members of the community. This signalling is transmitted through specific chemicals called autoinducers and this phenomena is called quorum sensing (QS) (Venturi 2006). QS is now widely recognized as an essential mechanism in many bacterial species. Most Gram negative bacteria regulate QS by production of autoinducer, N–acyl–L–homoserine lactones (AHLs) [reviewed in (Camilli and Bassler, 2006)]. This mechanism was first identified in a marine bacteria Vibrio fischeri which lives in symbiotic associations with many marine organisms and produces bioluminescence. The bioluminescence was seen only when the bacterial population–density was above certain threshold level. The bioluminescence was found to be associated with an increase in concentration of an autoinducer – (30–oxohexanoyl)–L–homoserine lactone (3–oxo–C6–HSL, 3) produced by V.fischeri cells [reviewed in (Dunlap 1999)]. It is now known that luxI/luxR circuit controls this mechanism. In bacteria LuxI, an autoinducer, synthase, catalyzes synthesis of AHL. The receptor protein, LuxR, usually remains inactive and is continuously degraded in the cells. When AHL binds with LuxR,it becomes activated and activated LuxR binds to DNA and starts transcription of the target genes. This QS mechanism has been found essential for exopolysaccharide biosynthesis, virulence, biofilm formation, and biofilm related antibiotic resistance [reviewed in (Rutherford and Bassler, 2012)].
In P. aeruginosatwo sets of luxI/luxR type systems have been identified. The lasI/lasR regulated QS system produces and senses N–(3–oxododecanoyl)– L–homoserine lactone (Passador et al., 1993), and the rhlI / rhlR–based system synthesizes and responds to N–butyryl homoserine lactone (Ochsner and Reiser, 1995). In addition P. aeruginosahas a third QS system, PQS which synthesizes 2–heptyl–3–hydroxy–4–quinolone, and regulates both the las and rhlQS systems (Diggle et al., 2006).
It has been shown that P. aeruginosabiofilm accumulates high concentrations of AHL molecules, i.e the concentration of N–(3–oxododecanoyl)–L–homoserine lactone recorded in biofilm was 630µM, whereas in effluent only 14 µM was found (Charlton et al., 2000). This high concentration is enough to initiate QS based responses in biofilm (Pearson et al., 1994). Many studies involving wild type and QS mutants have shown the effect of QS on biofilm development, architecture and exopolysaccharide biosynthesis (Bjarnsholt et al., 2005; De Kievit et al., 2001; Sakuragi and Kolter, 2007). Interestingly, a recent study has shown a link between c–di–GMP, QS and exopolysaccharide biosynthesis in P. aeruginosa. The study showed that AHL signals activate the protein TpbA which dephosphorylates TpbB, a PDE, and consequently leads to decreased c–di–GMP production (Ueda and Wood, 2009). This reduction in intracellular concentration of c–di–GMP, negatively impacts on exopolysaccharide production and leads to reduced biofilm formation.
THE IMPORTANT EXOPOLYSACCHARIDES FOR PSEUDOMONAS AERUGINOSA BIOFILM FORMATION
Three exopolysaccharides produced by P. aeruginosahave been identified as important for biofilm formation, architecture and function: Pel, Psl and alginate. Their detail is given below.
Molecular Structure of Pel
The structure of Pel has not been resolved yet. A comparison of sugar composition of the matrix from P. aeruginosastrain PA14 and Pel–deficient mutant revealed that the matrix of the Pel–deficient mutant contained less glucose than the Pel–producing strain (Friedman and Kolter, 2004). The study indirectly suggested that Pel is made of glucose. In contrast, a recent study provided an analysis of the total polymeric substances produced by PA14 and showed that the most abundant extracellular carbohydrates present in a PA14 culture were O–antigen lipopolysaccharide and glycerol–phosphorylated cyclic β–(1,3)–glucans. They did not detect any other glucose rich exopolysaccharide (Coulon et al., 2010).
Pel Biosynthesis
Pel synthesis and export machinery is encoded by pel operon which consist of seven genes: pelA, pelB, pelC, pelD pelE, pelF and pelG (Friedman and Kolter 2004) and all these genes have been shown to be essential for Pel biosynthesis (Vasseur et al., 2005). These pel genes are conserved in most Gram–negative bacteria and a study showed that a mutation in a homologue of pelG in Ralstonia solanacearum, ragG, resulted in defective in adherence of the bacteria to a solid surface (Vasseur et al., 2005). A little experimental work has been done around the structure and function of the proteins encoded by the pel genes. Only two proteins, PelC and PelD, have been experimentally examined to date (Kowalska et al., 2010; Lee et al., 2007; Ueda and Wood 2009; Vasseur et al., 2007). The function and structure of other proteins encoded by the pel operon have been suggested on the basis of sequence homology (Franklin et al., 2011; Keiski et al., 2010; Vasseur et al., 2005). Using these experimental evidences and bioinformatics analysis of the proteins encoded by the pel operon, hypothetical models for Pel exopolysaccharide biosynthesis has been proposed by different researchers (Franklin et al., 2011; Vasseur et al., 2007).
Biosynthesis of polysaccharide starts with polymerization of sugar molecules which are available as activated nucleotide–sugars (Rocchetta et al., 1999; Samuel and Reeves 2003; Whitfield and Roberts 1999). These activated sugars are usually synthesized by the enzymes encoded by genes from the polysaccharide biosynthesis operon. Interestingly, the pel operon lacks any such genes suggesting that precursors for Pel biosynthesis are taken from the main pool of nucleotide–sugars (Franklin et al., 2011). Recently it has been shown that AlgC, is involved in biosynthesis of glucose–1–phosphate, and algC negative mutants were not able to form Pel (Ma et al., 2012). It is possible that AlgC may be a common enzyme required for precursor synthesis for Pel, alginate and Psl. The nucleotide–sugars required for Pel biosynthesis are not known. On the basis of sugar composition analysis of exopolysaccharides produced by P. aeruginosaPAK strains, it is suggested that Pel is made up of glucose. This suggests that an activated–glucose may be used towards Pel biosynthesis. These activated sugars are transferred onto growing polymer by the enzymes called glycosyltransferases (GT). According to bioinformatics analysis, PelF is the only glycosyltransferase encoded by the pel operon (Franklin et al., 2011). On the basis of sequence similarities it is predicted that PelF belongs to the GT–4 family (according to the classification of CAZy(carbohydrate–active enzymes database) and members of this family are retaining glycosyltransferases (Coutinho et al., 2003). The retaining glycosyltransferases transfer sugar residues onto the acceptor molecule by retaining the anomeric configuration of the sugar as it was in the donor substrate. No transmembrane (TM) helices are predicted in PelF which is suggestive of a cytosolic subcellular localization. It is predicted that a cytosolic PelF synthesizes Pel on the cytosolic face of the inner membrane. It is not known how this newly synthesized polymer is transported across the plasma membrane. The pel operon contains only one glycosyltransferase, PelF which suggests that, like alginate, Pel is a homopolymer. Therefore, it is likely that Pel translocation occurs via a Wzy–independent pathway. In addition, the pel operon lacks a gene for an undecaprenyl–phosphate glycosylphosphotransferase which is required in undecaprenyl–phosphate dependent translocation of polysaccharide from the cytosol to the periplasm [reviewed in (Franklin et al., 2011)]. The newly polymerized Pel is likely to be translocated to the periplasm by PelD or PelE or PelG, because all of these proteins are predicted to be located in the inner membrane and contain multiple TM domains [reviewed in (Franklin et al., 2011)]. PelD binds with c–di–GMP and regulates the Pel biosynthesis (Lee et al., 2007). The role of c–di–GMP can be linked with Pel production because when the intracellular concentration of c–di–GMP was manipulated by deletion of tyrosine phosphatase, TpbA, the amount of Pel production and pellicle formation was also changed (Ueda and Wood, 2009). PelG is predicted to be an inner membrane protein which contains 12 TM domains (Krogh et al., 2001). On the basis of structural homology, PelG has been shown to be similar to NorM from V. cholerae (PDB ID: 3MKU). NorM belongs to the multidrug and toxic compound extrusion (MATE) family of proteins (He et al., 2010). Members of this family of proteins are involved in extrusion of molecules across the inner membrane. PelG could be involved in export of Pel polysaccharide from the cytoplasmic face of the inner membrane to the periplasm. PelE, the third inner membrane protein, is predicted to have two transmembrane domains which orient the protein in such a way that the C–terminus becomes localized to the periplasm. The C–terminus of the protein is predicted to contain 4–6 copies of tetratricopeptide–like repeats (TPR), the protein–protein interaction motif. Previously it has been shown TPR motifs are involved in the assembly of large protein complexes (Blatch and Lassle 1999; D'Andrea and Regan, 2003), therefore, PelE, may be involved in interaction with other TPR–containing proteins in the periplasm. The homology modelling suggests that the N–terminus of PelB forms a domain which has similarity with TPR–containing anaphase–promoting complex/cyclosome subunit Cdc 16/Cut9 [PDB ID: 2XPI; (Zhang et al., 2010)] and is predicted to contain TPR motifs. It can be proposed that the C–terminus of PelE and N–terminus of PelB are involved in protein interactions to make a scaffold. PelC has been proposed as a lipoprotein in which Cystein–19 is lipidated and contains a C–terminal amphipathic α–helix which is inserted in the outer membrane (OM) in a Wza–like manner. This C–terminus domain has been shown to be essential for Pel biosynthesis. PelA is a large 105 kDa protein in which an N–terminal domain is structurally similar to glycoside hydrolases (PDB ID: 2AAM) and a C–terminal domain shows similarity to carbohydrate esterase [PDB ID: 2VYO; (Urch et al., 2009)]. The glycosidic hydrolase activity of PelA suggests that PelA may play a role similar to AlgL. AlgL hydrolyses alginate which escapes into the periplasm (Bakkevig et al., 2005). Therefore, it is possible that any Pel that escapes into the periplasm is hydrolyzed by PelA.
Pel biosynthesis can be summarized as PelF polymerizing a homopolymer of glucose which is transferred from the cytoplasm to the periplasm by PelE /PelD/ PelG.
Molecular Structure of Psl
Previous studies suggests that Psl is composed of galactose, D–mannose, D–glucose, and L–rhamnose (Ma et al., 2007; Ma et al., 2009) Recently the molecular structure of Psl is revealed (Byrd et al., 2009). The study showed that Psl exopolysaccharide is composed of pentasaccharides repeating units and each unit is composed of three D–mannose, one D–glucose, and one L–rhamnose.
Psl Biosynthesis
Biosynthesis of Psl in P. aeruginosais under the control of thepsl operon which contains 15 co–transcribed genes: pslA–pslO(Friedman and Kolter 2004). Interestingly, only 11 of the 15 gene products of thepsl operon were found to be essential for Psl biosynthesis. The product of pslB has redundant function with WbpW, an enzyme which has similar activity to that predicted for PslB (Byrd et al., 2009). Deletion of the last three genes of the operon, pslMNO, has shown no effect on the ability of P. aeruginosato produce Psl (Byrd et al., 2009).
Although the Psl biosynthesis mechanism has not been established experimentally, the repeating unit structure of Psl and bioinformatics analysis of the proteins encoded by the psl operon suggest that Psl biosynthesis bears a resemblance to the undecaprenyl diphosphate–based biosynthesis of E. coli group–1 capsular and extracellular polysaccharides. Therefore the Psl biosynthesis pathway resembles a Wzy–dependent pathway. This hypothesis is further strengthened by the finding that three proteins encoded by the corresponding genes of the psl operon PslA, PslD and PslE show sequence similarity with WbaP, Wza, and Wzc, respectively(Franklin et al., 2011). The role of gene products of psl operon is presented in a recent review (Franklin et al., 2011).
The detailed account of alginate is reviewed elsewhere in the review article (Hay et al., 2010)
ROLE OF EXOPOLYSACCHARIDES IN BIOFILM
Role of Pel in Biofilm
Pel is synthesized in most P. aeruginosastrains but strains with intact psl loci depend more on Psl contributing to biofilm formation and architecture. However, P aeruginosa PA14 relies mainly on the Pel exopolysaccharide because it lacks Psl (Friedman and Kolter, 2004). This strain produces biofilm at the air–liquid interface (A–L) and it was observed that a pel transposon insertion mutant failed to produce A–L biofilm (Friedman and Kolter 2004). This finding led to further detailed investigation of the role of Pel in biofilm formation. Pel has been shown to be required for adherence to abiotic surfaces especially at the later stages of the biofilm in PA14 (Colvin et al., 2011). Recent studies have suggested a redundant role for Pel in P aeruginosaPAO1 biofilm, especially in the presence of Psl. However, in the absence of Psl, Pel has a clearer role in biofilm formation (Colvin et al., 2012). In contrast, a recent study has demonstrated a distinct role for Pel in biofilm formation and suggested that, in addition to Psl, Pel was required for structural stability of mature biofilms (Yang et al., 2011).
Pel has also been identified as an extracellular matrix component which mitigates the effects of antibiotics and may play a vital role in protection of the bacteria (Colvin et al., 2011). Studies demonstrated that Pel–producing PA14 strains showed increased resistance to aminoglycoside antibiotics when compared to Pel–deficient strains (Colvin et al., 2011). In contrast, other authors have reported contradicting results which showed that Pel–deficient mutants were more resistant to the aminoglycoside, tobramycin when compared to wild type PA14 strains (Khan et al., 2010). The two studies used different methods and conditions to assess antibiotic resistance which might have affected the results of their experiments. (Colvin et al., 2011) grew biofilms on polycarbonate filters for 48 hours and then the filter was transferred to solid medium containing respective antibiotics and incubated for 24 hours.Viable bacterial cells were counted to see the effect of antibiotics. It was found that absence of Pel renders PA14 cells more susceptible to aminoglycosides when grown as biofilm. Whereas, (Khan et al., 2010) grew cells in 96–well microtiter plates in liquid medium for 24 hours. These one day old biofilms were washed and treated with respective antibiotics for 24 hours. It was found that in absence of Pel PA14 cells were more resistant to aminoglycosides. This difference of media conditions and duration of growth might have affected the exopolysaccharide production and distribution in biofilm formation.Owing to the significance of Pel in antibiotic resistance and architecture of the biofilm, it is imperative to further investigate its regulation, biosynthesis and role in P. aeruginosabiofilm.
Role of Psl in Biofilm
Psl is found in two forms: a larger form associated with cells and a smaller soluble form. It is not yet clear how the smaller form of Psl is produced. Possibly, it may be produced by cleavage or breakdown of the longer cell–associated Psl. Alternatively, there could be another unrecognized mechanism to generate this form of Psl. The long cell–associated form of Psl forms blebs of Psl on the cell surface which can be observed by scanning electron microscopy (SEM) (Byrd et al., 2010). Psl has been shown to be important for initial attachment of non–motile cells to both biotic and abiotic surfaces (Ma et al., 2006; Byrd et al., 2009; Byrd et al., 2010). Psl aids structural stability of the biofilm and is required for the typical architecture of the biofilm (Ma et al., 2009). Psl also plays an important role in protection of bacterial cells from innate immune effectors, complement and neutrophils (Mishra et al., 2012). Recently it has been shown that Psl also stimulates the production of c–di–GMP in surrounding bacterial cells suggesting a regulatory role of Psl in biofilm formation and maintenance (Irie et al., 2012).
Role of Alginate in Biofilm
Most of the early studies investigating P. aeruginosabiofilms were conducted using mucoid (alginate overproducing) strains (Ohman and Chakrabarty, 1982; Pedersen et al., 1992; Sa–Correia et al., 1987). Mucoid strains isolated from chronically infected lungs of cystic fibrosis patients were able to produce copious amount of alginate when grown on agar plates in the laboratory. Mucoid colonies, encased in matrix made of alginate, appear more slimy when compared to non–mucoid strains (Hogardt and Heesemann, 2010). Alginate has been shown to be a very important matrix component for retention of water due to its gel–forming ability and protects the bacterial cells from the harmful effects of desiccation. Although direct evidence of the amount of water retained by alginate is not known, a similar exopolysaccharide, hyaluronic acid, can bind up to 1 kg of water per gram of the polysaccharide (Sutherland, 2001). The degree of acetylation of alginate also plays an important role in its ability to retain water. High degrees of O – acetylation of the residues present in alginate increase its solubility by reducing the chance of interaction between alginate and cations. Alginates without O–acetyl residues show increased binding of divalent cations and decreased polysaccharide solubility (Sutherland, 2001). Alginate protects bacterial cells from the host immune response and antimicrobial drugs (Simpson et al., 1988; Simpson et al., 1989; Simpson et al., 1993). Most of the host immune cells such as neutrophils and activated macrophages kill bacterial cells by releasing free radicals. Alginate can protect bacterial cells by scavenging the free radicals (Simpson et al., 1988; Simpson et al., 1989).
Role of Exopolysaccharides in the Architecture of the Biofilm
The production and quantity of exopolysaccharides influences the architecture of biofilm. Studies show that mutants lacking exopolysaccharides produce biofilms which are structurally different from those produced by the wild type strains. For example, V. cholerae deficient in exopolysaccharides (Watnick and Kolter, 1999) and E. coli lacking colanic acid (Danese et al., 2000) failed to form 3D biofilm architecture. The fruiting body–like biofilms formed by wild type B. subtilis were absent in the mutants lacking exopolysaccharides and the secreted protein TasA (Branda et al., 2006). Less is known about the effects of polysaccharides on the architecture of P. aeruginosabiofilms. Some studies have been carried out to investigate the role of alginate in biofilm architecture (Franklin and Ohman 1993; Hay et al., 2009; Wozniak et al., 2003). Although it has been suggested that alginate is not necessary for P. aeruginosabiofilm formation (Wozniak et al., 2003), however, biofilm architecture is influenced in the presence of alginate. Some recent studies have shown the impact of Psl in biofilm architecture (Overhage et al., 2005; Ma et al., 2006; Starkey et al., 2009). Further investigations are required to decipher the role of Pel in the architecture of P. aeruginosabiofilms.
REFERENCES
Agladze K, Wang X, Romeo T (2005). Spatial periodicity of Escherichia coli K–12 biofilm microstructure initiates during a reversible, polar attachment phase of development and requires the polysaccharide adhesin PGA. J. Bacteriol. 187 (24): 8237 – 8246.
http://dx.doi.org/10.1128/JB.187.24.8237-8246.2005
PMid:16321928 PMCid:PMC1317006
Allegrucci M, Hu FZ, Shen K, Hayes J, Ehrlich GD, Post JC, Sauer K (2006). Phenotypic characterization of Streptococcus pneumoniae biofilm development. J. Bacteriol. 188 (7): 2325 – 2335.
http://dx.doi.org/10.1128/JB.188.7.2325-2335.2006
PMid:16547018 PMCid:PMC1428403
Allesen–Holm M, Barken KB, Yang L, Klausen M, Webb JS, Kjelleberg S, Molin S, Givskov M, Tolker–Nielsen T (2006). A characterization of DNA release in Pseudomonas aeruginosa cultures and biofilms. Mol. Microbiol. 59 (4): 1114 – 1128.
http://dx.doi.org/10.1111/j.1365-2958.2005.05008.x
PMid:16430688
Arora SK, Ritchings BW, Almira EC, Lory S, Ramphal R (1998). The Pseudomonas aeruginosa flagellar cap protein, FliD, is responsible for mucin adhesion. Infect. Immun. 66(3): 1000 – 1007.
PMid:9488388 PMCid:PMC108008
Bakkevig K, Sletta H, Gimmestad M, Aune R, Ertesvag H, Degnes K, Christensen BE, Ellingsen TE, Valla S (2005). Role of the Pseudomonas fluorescens alginate lyase (AlgL) in clearing the periplasm of alginates not exported to the extracellular environment. J. Bacteriol. 187 (24): 8375 – 8384.
http://dx.doi.org/10.1128/JB.187.24.8375-8384.2005
PMid:16321942 PMCid:PMC1317005
Banin E, Vasil ML, Greenberg EP (2005). Iron and Pseudomonas aeruginosa biofilm formation. Proc. Natl. Acad. Sci. USA. 102 (31): 11076 – 11081.
http://dx.doi.org/10.1073/pnas.0504266102
PMid:16043697 PMCid:PMC1182440
Baraquet C, Murakami K, Parsek MR, Harwood CS (2012). The FleQ protein from Pseudomonas aeruginosa functions as both a repressor and an activator to control gene expression from the pel operon promoter in response to c–di–GMP. Nucleic Acids Res. 40 (15): 7207 – 7218.
http://dx.doi.org/10.1093/nar/gks384
PMid:22581773 PMCid:PMC3424551
Barken KB, Pamp SJ, Yang L, Gjermansen M, Bertrand JJ, Klausen M, Givskov M, Whitchurch CB, Engel JN, Tolker–Nielsen T (2008). Roles of type IV pili, flagellum–mediated motility and extracellular DNA in the formation of mature multicellular structures in Pseudomonas aeruginosa biofilms. Environ. Microbiol. 10(9): 2331 – 2343.
http://dx.doi.org/10.1111/j.1462-2920.2008.01658.x
PMid:18485000
Barraud N, Schleheck D, Klebensberger J, Webb JS, Hassett DJ, Rice SA, Kjelleberg S (2009). Nitric oxide signaling in Pseudomonas aeruginosa biofilms mediates phosphodiesterase activity, decreased cyclic di–GMP levels, and enhanced dispersal. J. Bacteriol. 191 (23): 7333 – 7342.
http://dx.doi.org/10.1128/JB.00975-09
PMid:19801410 PMCid:PMC2786556
Bendinger B, Rijnaarts HH, Altendorf K, Zehnder AJ (1993). Physicochemical cell surface and adhesive properties of coryneform bacteria related to the presence and chain length of mycolic acids. Appl. Environ. Microbiol. 59(11): 3973 – 3977.
PMid:16349100 PMCid:PMC182562
Bjarnsholt T, Jensen PO, Burmolle M, Hentzer M, Haagensen JA, Hougen HP, Calum H, Madsen KG, Moser C, Molin S, Hoiby N, Givskov M (2005). Pseudomonas aeruginosa tolerance to tobramycin, hydrogen peroxide, polymorphonuclear leukocytes is quorum–sensing dependent. Microbiol. 151: 373 – 383.
http://dx.doi.org/10.1099/mic.0.27463-0
PMid:15699188
Bjarnsholt T, Jensen PØ, Fiandaca MJ, Pedersen J, Hansen CR, Andersen CB, Pressler T, Givskov M, Høiby N (2009). Pseudomonas aeruginosa biofilms in the respiratory tract of cystic fibrosis patients. Pediatric Pulmonol. 44 (6): 547 – 558.
http://dx.doi.org/10.1002/ppul.21011
PMid:19418571
Blatch GL, Lassle M (1999). The tetratricopeptide repeat: a structural motif mediating protein–protein interactions. BioEssays. 21 (11): 932 – 939.
http://dx.doi.org/10.1002/(SICI)1521-1878(199911)21:11<932::AID-BIES5>3.3.CO;2-E
http://dx.doi.org/10.1002/(SICI)1521-1878(199911)21:11<932::AID-BIES5>3.0.CO;2-N
Borlee BR, Goldman AD, Murakami K, Samudrala R, Wozniak DJ, Parsek MR (2010). Pseudomonas aeruginosa uses a cyclic–di–GMP–regulated adhesin to reinforce the biofilm extracellular matrix. Mol. Microbiol. 75 (4): 827 – 842.
http://dx.doi.org/10.1111/j.1365-2958.2009.06991.x
PMid:20088866 PMCid:PMC2847200
Branda SS, Chu F, Kearns DB, Losick R, Kolter R (2006). A major protein component of the Bacillus subtilis biofilm matrix. Mol. Microbiol. 59 (4): 1229 – 1238.
http://dx.doi.org/10.1111/j.1365-2958.2005.05020.x
PMid:16430696
Branda SS, Vik S, Friedman L, Kolter R (2005). Biofilms: the matrix revisited. Trends Microbiol. 13 (1): 20 – 26.
http://dx.doi.org/10.1016/j.tim.2004.11.006
PMid:15639628
Bucior I, Pielage JF, Engel JN (2012). Pseudomonas aeruginosa pili and flagella mediate distinct binding and signaling events at the apical and basolateral surface of airway epithelium. PLoS Pathog. 8(4): e1002616.
http://dx.doi.org/10.1371/journal.ppat.1002616
PMid:22496644 PMCid:PMC3320588
Bullitt E, Makowski L (1995). Structural polymorphism of bacterial adhesion pili. Nat. 373 (6510): 164 – 167.
http://dx.doi.org/10.1038/373164a0
PMid:7816100
Byrd MS, Pang B, Mishra M, Swords WE, Wozniak DJ (2010). The Pseudomonas aeruginosa exopolysaccharide Psl facilitates surface adherence and NF–kappaB activation in A549 cells. mBio. 1(3): e00140-10.
Byrd MS, Sadovskaya I, Vinogradov E, Lu H, Sprinkle AB, Richardson SH, Ma L, Ralston B, Parsek MR, Anderson EM, Lam JS, Wozniak DJ (2009). Genetic and biochemical analyses of the Pseudomonas aeruginosa Psl exopolysaccharide reveal overlapping roles for polysaccharide synthesis enzymes in Psl and LPS production. Mol. Microbiol. 73: 622 – 638.
http://dx.doi.org/10.1111/j.1365-2958.2009.06795.x
PMid:19659934
Caiazza NC, Merritt JH, Brothers KM, O'Toole GA (2007). Inverse regulation of biofilm formation and swarming motility by Pseudomonas aeruginosa PA14. J. Bacteriol. 189 (9): 3603 – 3612.
http://dx.doi.org/10.1128/JB.01685-06
PMid:17337585 PMCid:PMC1855903
Caiazza NC, O'Toole GA (2004). SadB Is Required for the Transition from Reversible to Irreversible Attachment during Biofilm Formation by Pseudomonas aeruginosa PA14. J. Bacteriol. 186 (14): 4476 – 4485.
http://dx.doi.org/10.1128/JB.186.14.4476-4485.2004
PMid:15231779 PMCid:PMC438627
Camilli A, Bassler BL (2006). Bacterial small–molecule signaling pathways. Sci. 311 (5764): 1113 – 1116.
http://dx.doi.org/10.1126/science.1121357
PMid:16497924 PMCid:PMC2776824
Characklis WG (1973). Attached microbial growths—II. Frictional resistance due to microbial slimes. Water Res. 7 (9): 1249 – 1258.
http://dx.doi.org/10.1016/0043-1354(73)90002-X
Characklis WG GM, KC Marshall (1990). Physiological ecology in biofilm systems. In: K. M. Characklis WG (eds), Biofilms. John Wiley & Sons, New York. 341 – 394.
Charlton TS, de Nys R, Netting A, Kumar N, Hentzer M, Givskov M, Kjelleberg S (2000). A novel and sensitive method for the quantification of N–3–oxoacyl homoserine lactones using gas chromatography–mass spectrometry: application to a model bacterial biofilm. Environ.Microbiol. 2(5): 530 – 541.
http://dx.doi.org/10.1046/j.1462-2920.2000.00136.x
PMid:11233161
Choy WK, Zhou L, Syn CK, Zhang LH, Swarup S (2004). MorA defines a new class of regulators affecting flagellar development and biofilm formation in diverse Pseudomonas species. J. Bacteriol. 186(21): 7221 – 7228.
http://dx.doi.org/10.1128/JB.186.21.7221-7228.2004
PMid:15489433 PMCid:PMC523210
Christen M, Christen B, Folcher M, Schauerte A, Jenal U (2005). Identification and Characterization of a Cyclic di–GMP–specific Phosphodiesterase and Its Allosteric Control by GTP. J. Biol. Chem. 280(35): 30829 – 30837.
http://dx.doi.org/10.1074/jbc.M504429200
PMid:15994307
Colombini S, Merchant SR, Hosgood G (2000). Microbial flora and antimicrobial susceptibility patterns from dogs with otitis media. Vet. Dermatol. 11(4): 235 – 239.
http://dx.doi.org/10.1046/j.1365-3164.2000.00191.x
Colvin KM, Gordon VD, Murakami K, Borlee BR, Wozniak DJ, Wong GC, Parsek MR (2011). The pel polysaccharide can serve a structural and protective role in the biofilm matrix of Pseudomonas aeruginosa . PLoS Pathol. 7(1): e1001264.
http://dx.doi.org/10.1371/journal.ppat.1001264
PMid:21298031 PMCid:PMC3029257
Colvin KM, Gordon VD, Murakami K, Borlee BR, Wozniak DJ, Wong GC, Parsek MR (2011). The pel polysaccharide can serve a structural and protective role in the biofilm matrix of Pseudomonas aeruginosa . PLoS Pathol. 7(1): e1001264.
http://dx.doi.org/10.1371/journal.ppat.1001264
PMid:21298031 PMCid:PMC3029257
Colvin KM, Irie Y, Tart CS, Urbano R, Whitney JC, Ryder C, Howell PL, Wozniak DJ, Parsek MR (2012). The Pel and Psl polysaccharides provide Pseudomonas aeruginosa structural redundancy within the biofilm matrix. Environ.Microbiol. 14(8): 1913 – 1928.
http://dx.doi.org/10.1111/j.1462-2920.2011.02657.x
PMid:22176658 PMCid:PMC3840794
Corpe W (1980). Microbial surface components involved in adsorption of microorganisms onto surfaces. In: K. M. Bitton G (eds), Adsorption of microorganisms to surfaces. John Wiley & Sons, New York. 105–144.
Costerton JW, Geesey GG, Cheng KJ (1978). How bacteria stick. Sci. Am. 238(1): 86 – 95.
http://dx.doi.org/10.1038/scientificamerican0178-86
PMid:635520
Costerton JW, Lewandowski Z, Caldwell DE, Korber DR, Lappin–Scott HM (1995). Microbial biofilms. Annu. Rev. Microbiol. 49: 711 – 745.
http://dx.doi.org/10.1017/CBO9780511525353.002
Costerton JW, Stewart PS, Greenberg EP (1999). Bacterial biofilms: a common cause of persistent infections. Science. 284(5418): 1318 – 1322.
http://dx.doi.org/10.1126/science.284.5418.1318
PMid:10334980
Coulon C, Vinogradov E, Filloux A, Sadovskaya I (2010). Chemical analysis of cellular and extracellular carbohydrates of a biofilm–forming strain Pseudomonas aeruginosa PA14. PLoS ONE. 5 (12): e14220.
http://dx.doi.org/10.1371/journal.pone.0014220
PMid:21151973 PMCid:PMC2997053
Coutinho PM, Deleury E, Davies GJ, Henrissat B (2003). An evolving hierarchical family classification for glycosyltransferases. J. Mol. Biol. 328 (2): 307 – 317.
http://dx.doi.org/10.1016/S0022-2836(03)00307-3
D'Andrea LD, Regan L (2003). TPR proteins: the versatile helix. Trends Biochem. Sci. 28 (12): 655 – 662.
http://dx.doi.org/10.1016/j.tibs.2003.10.007
PMid:14659697
Danese PN, Pratt LA, Kolter R (2000). Exopolysaccharide production is required for development of Escherichia coli K–12 biofilm architecture. J. Bacteriol. 182 (12): 3593 – 3596.
http://dx.doi.org/10.1128/JB.182.12.3593-3596.2000
PMid:10852895 PMCid:PMC101973
Darby C, Hsu JW, Ghori N, Falkow S (2002). Caenorhabditis elegans: plague bacteria biofilm blocks food intake. Nat. 417 (6886): 243 – 244.
http://dx.doi.org/10.1038/417243a
PMid:12015591
De Kievit TR, Gillis R, Marx S, Brown C, Iglewski BH (2001). Quorum–sensing genes in Pseudomonas aeruginosa biofilms: their role and expression patterns. Appl. Environ. Microbiol. 67 (4): 1865 – 1873.
http://dx.doi.org/10.1128/AEM.67.4.1865-1873.2001
PMid:11282644 PMCid:PMC92808
DeVault JD, Kimbara K, Chakrabarty AM (1990). Pulmonary dehydration and infection in cystic fibrosis: evidence that ethanol activates alginate gene expression and induction of mucoidy in Pseudomonas aeruginosa . Mol. Microbiol. 4 (5): 737 – 745.
http://dx.doi.org/10.1111/j.1365-2958.1990.tb00644.x
PMid:2167423
Diggle SP, Cornelis P, Williams P, Camara M (2006). 4–quinolone signalling in Pseudomonas aeruginosa : old molecules, new perspectives. Int. J. Med. Microbiol. 296 (2–3): 83 – 91.
http://dx.doi.org/10.1016/j.ijmm.2006.01.038
PMid:16483840
Diggle SP, Stacey RE, Dodd C, Camara M, Williams P, Winzer K (2006). The galactophilic lectin, LecA, contributes to biofilm development in Pseudomonas aeruginosa . Environ.Microbiol. 8 (6): 1095 – 1104.
http://dx.doi.org/10.1111/j.1462-2920.2006.001001.x
PMid:16689730
Donlan RM (2002). Biofilms: microbial life on surfaces. Emerg. Infect. Dis. 8 (9): 881 – 890.
http://dx.doi.org/10.3201/eid0809.020063
PMid:12194761 PMCid:PMC2732559
Dunlap PV (1999). Quorum regulation of luminescence in Vibrio fischeri. J. Mol. Microbiol. Biotechnol. 1 (1): 5 – 12.
PMid:10941779
Flemming HC, Neu TR, Wozniak DJ (2007). The EPS matrix: the "house of biofilm cells". J. Bacteriol. 189 (22): 7945 – 7947.
http://dx.doi.org/10.1128/JB.00858-07
PMid:17675377 PMCid:PMC2168682
Flemming H–C, Wingender GJ, Mayer C (2000). Physico–chemical properties of biofilms. In: L. Evans (eds), Biofilms: recent advances in their study and control. Harwood Academic Publishers, Amsterdam. 19 – 34.
Flemming HC, Wingender J (2010). The biofilm matrix. Nat. Rev. Microbiol. 8 (9): 623 – 633.
PMid:20676145
Fletcher M, Loeb GI (1979). Influence of substratum characteristics on the attachment of a marine pseudomonad to solid surfaces. Appl. Environ. Microbiol. 37 (1): 67 – 72.
PMid:16345338 PMCid:PMC243402
Franklin MJ, Nivens DE, Weadge JT, Howell PL (2011). Biosynthesis of the Pseudomonas aeruginosa extracellular polysaccharides, alginate, Pel, and Psl. Front. Microbio. 2: 167.
http://dx.doi.org/10.3389/fmicb.2011.00167
PMid:21991261 PMCid:PMC3159412
Franklin MJ, Ohman DE (1993). Identification of algF in the alginate biosynthetic gene cluster of Pseudomonas aeruginosa which is required for alginate acetylation. J. Bacteriol. 175 (16): 5057 – 5065.
PMid:8394313 PMCid:PMC204972
Friedman L, Kolter R (2004). Genes involved in matrix formation in Pseudomonas aeruginosa PA14 biofilms. Mol. Microbiol. 51 (3): 675 – 690.
http://dx.doi.org/10.1046/j.1365-2958.2003.03877.x
PMid:14731271
Friedman L, Kolter R (2004). Two genetic loci produce distinct carbohydrate–rich structural components of the Pseudomonas aeruginosa biofilm matrix. J. Bacteriol. 186 (14): 4457 – 4465.
http://dx.doi.org/10.1128/JB.186.14.4457-4465.2004
PMid:15231777 PMCid:PMC438632
Fuqua C (2010). Passing the baton between laps: adhesion and cohesion in Pseudomonas putida biofilms. Mol. Microbiol. 77 (3): 533 – 536.
http://dx.doi.org/10.1111/j.1365-2958.2010.07250.x
PMid:20545855 PMCid:PMC2909372
Galperin MY, Nikolskaya AN, Koonin EV (2001). Novel domains of the prokaryotic two–component signal transduction systems. FEMS Microbiol. Lett. 203 (1): 11 – 21.
http://dx.doi.org/10.1111/j.1574-6968.2001.tb10814.x
PMid:11557134
Geesey GG, Richardson WT, Yeomans HG, Irvin RT, Costerton JW (1977). Microscopic examination of natural sessile bacterial populations from an alpine stream. Can. J. Microbiol. 23 (12): 1733 – 1736.
http://dx.doi.org/10.1139/m77-249
PMid:340020
Gloag ES, Turnbull L, Huang A, Vallotton P, Wang H, Nolan LM, Mililli L, Hunt C, Lu J, Osvath SR, Monahan LG, Cavaliere R, Charles IG, Wand MP, Gee ML, Prabhakar R, Whitchurch CB (2013). Self–organization of bacterial biofilms is facilitated by extracellular DNA. Proceedings of the National Academy of Sciences.
http://dx.doi.org/10.1073/pnas.1218898110
PMid:23798445 PMCid:PMC3710876
Götz F (2002). Staphylococcus and biofilms. Mol. Microbiol. 43 (6): 1367 – 1378.
http://dx.doi.org/10.1046/j.1365-2958.2002.02827.x
PMid:11952892
Goulter RM, Gentle IR, Dykes GA (2009). Issues in determining factors influencing bacterial attachment: a review using the attachment of Escherichia coli to abiotic surfaces as an example. Lett. Appl. Microbiol. 49 (1): 1 – 7.
http://dx.doi.org/10.1111/j.1472-765X.2009.02591.x
PMid:19291206
Hamilton W (1987). Biofilm: microbial interaction and metabolic activities. Ecology of microbial communities. T. G. Fletcher M, JG Jones. Cambridge, Cambridge University Press 361 – 387.
Harmsen M, Yang LA, Pamp SJ, Tolker–Nielsen T (2010). An update on Pseudomonas aeruginosa biofilm formation, tolerance and dispersal. FEMS Immunol. Med. Microbiol. 59 (3): 253 – 268.
PMid:20497222
Hay ID, Gatland K, Campisano A, Jordens JZ, Rehm BH (2009). Impact of alginate overproduction on attachment and biofilm architecture of a supermucoid Pseudomonas aeruginosa strain. Appl. Environ. Microbiol. 75 (18): 6022 – 6025.
http://dx.doi.org/10.1128/AEM.01078-09
PMid:19648373 PMCid:PMC2747843
Hay ID, Remminghorst U, Rehm BH (2009). MucR, a novel membrane–associated regulator of alginate biosynthesis in Pseudomonas aeruginosa . Appl. Environ. Microbiol. 75 (4): 1110 – 1120.
http://dx.doi.org/10.1128/AEM.02416-08
PMid:19088322 PMCid:PMC2643583
Hay ID, Ur Rehman Z, Ghafoor A, Rehm BHA (2010). Bacterial biosynthesis of alginates. J. Chem. Technol. Biotechnol. 85 (6): 752 – 759.
http://dx.doi.org/10.1002/jctb.2372
He X, Szewczyk P, Karyakin A, Evin M, Hong WX, Zhang Q, Chang G (2010). Structure of a cation–bound multidrug and toxic compound extrusion transporter. Nature. 467 (7318): 991 – 994.
http://dx.doi.org/10.1038/nature09408
PMid:20861838 PMCid:PMC3152480
Hendrickson El Fau – Plotnikova J, Plotnikova J Fau – Mahajan–Miklos S, Mahajan–Miklos S Fau – Rahme LG, Rahme Lg Fau – Ausubel FM, FM A (2001). Differential roles of the Pseudomonas aeruginosa PA14 rpoN gene in pathogenicity in plants, nematodes, insects, and mice. J. Bacteriol. 183 (24): 7126 – 7134.
http://dx.doi.org/10.1128/JB.183.24.7126-7134.2001
PMid:11717271 PMCid:PMC95561
Hengge R (2009). Principles of c–di–GMP signalling in bacteria. Nat. Rev. Microbiol. 7 (4): 263 – 273.
http://dx.doi.org/10.1038/nrmicro2109
PMid:19287449
Heukelekian H, Heller A (1940). Relation between food concentration and surface for bacterial growth. J. Bacteriol. 40 (4): 547 – 558.
PMid:16560368 PMCid:PMC374658
Hinsa SM, Espinosa–Urgel M, Ramos JL, O'Toole GA (2003). Transition from reversible to irreversible attachment during biofilm formation by Pseudomonas fluorescens WCS365 requires an ABC transporter and a large secreted protein. Mol. Microbiol. 49 (4): 905 – 918.
http://dx.doi.org/10.1046/j.1365-2958.2003.03615.x
PMid:12890017
Hogardt M, Heesemann J (2010). Adaptation of Pseudomonas aeruginosa during persistence in the cystic fibrosis lung. Int. J. Med. Microbiol. 300 (8): 557 – 562.
http://dx.doi.org/10.1016/j.ijmm.2010.08.008
PMid:20943439
Huang B, Whitchurch CB, Mattick JS (2003). FimX, a multidomain protein connecting environmental signals to twitching motility in Pseudomonas aeruginosa . J. Bacteriol. 185 (24): 7068 – 7076.
http://dx.doi.org/10.1128/JB.185.24.7068-7076.2003
PMid:14645265 PMCid:PMC296245
Humphrey B, Kjelleberg S, Marshall KC (1983). Responses of marine bacteria under starvation conditions at a solid–water interface. Appl. Environ. Microbiol. 45 (1): 43 – 47.
PMid:16346180 PMCid:PMC242228
Irie Y, Borlee BR, O'Connor JR, Hill PJ, Harwood CS, Wozniak DJ, Parsek MR (2012). Self–produced exopolysaccharide is a signal that stimulates biofilm formation in Pseudomonas aeruginosa . Proc. Natl. Acad. Sci. USA. 109 (50): 20632 – 20636.
http://dx.doi.org/10.1073/pnas.1217993109
PMid:23175784 PMCid:PMC3528562
Janakiraman RS, Brun YV (1999). Cell cycle control of a holdfast attachment gene in Caulobacter crescentus. J. Bacteriol. 181 (4): 1118 – 1125.
PMid:9973336 PMCid:PMC93487
Johansson EM, Crusz SA, Kolomiets E, Buts L, Kadam RU, Cacciarini M, Bartels KM, Diggle SP, Camara M, Williams P, Loris R, Nativi C, Rosenau F, Jaeger KE, Darbre T, Reymond JL (2008). Inhibition and dispersion of Pseudomonas aeruginosa biofilms by glycopeptide dendrimers targeting the fucose–specific lectin LecB. Chem. Biol. 15 (12): 1249 – 1257.
http://dx.doi.org/10.1016/j.chembiol.2008.10.009
PMid:19101469
Jones HC, Roth IL, Sanders WM, 3rd (1969). Electron microscopic study of a slime layer. J. Bacteriol. 99 (1): 316 – 325.
PMid:5802613 PMCid:PMC250005
Jurcisek JA, Bakaletz LO (2007). Biofilms formed by nontypeable Haemophilus influenzae in vivo contain both double–stranded DNA and type IV pilin protein. J. Bacteriol. 189 (10): 3868 – 3875.
http://dx.doi.org/10.1128/JB.01935-06
PMid:17322318 PMCid:PMC1913342
Kadurugamuwa JL, Beveridge TJ (1995). Virulence factors are released from Pseudomonas aeruginosa in association with membrane vesicles during normal growth and exposure to gentamicin: a novel mechanism of enzyme secretion. J. Bacteriol. 177 (14): 3998 – 4008.
PMid:7608073 PMCid:PMC177130
Kanbe M, Shibata S, Umino Y, Jenal U, Aizawa SI (2005). Protease susceptibility of the Caulobacter crescentus flagellar hook–basal body: a possible mechanism of flagellar ejection during cell differentiation. Microbiol. 151 (2): 433 – 438.
http://dx.doi.org/10.1099/mic.0.27386-0
PMid:15699192
Keiski CL, Harwich M, Jain S, Neculai AM, Yip P, Robinson H, Whitney JC, Riley L, Burrows LL, Ohman DE, Howell PL (2010). AlgK is a TPR–containing protein and the periplasmic component of a novel exopolysaccharide secretin. Structure. 18 (2): 265 – 273.
http://dx.doi.org/10.1016/j.str.2009.11.015
PMid:20159471 PMCid:PMC2857933
Khan W, Bernier SP, Kuchma SL, Hammond JH, Hasan F, O'Toole GA (2010). Aminoglycoside resistance of Pseudomonas aeruginosa biofilms modulated by extracellular polysaccharide. Int. Microbiol. 13 (4): 207 – 212.
PMid:21404215 PMCid:PMC3721063
Kierek K, Watnick PI (2003). Environmental determinants of Vibrio choleraebiofilm development. Appl. Environ. Microbiol. 69 (9): 5079 – 5088.
http://dx.doi.org/10.1128/AEM.69.9.5079-5088.2003
PMid:12957889 PMCid:PMC194957
Kirov SM, Webb JS, O'May C Y, Reid DW, Woo JK, Rice SA, Kjelleberg S (2007). Biofilm differentiation and dispersal in mucoid Pseudomonas aeruginosa isolates from patients with cystic fibrosis. Microbiol. 153: 3264 – 3274.
http://dx.doi.org/10.1099/mic.0.2007/009092-0
PMid:17906126
Klausen M, Heydorn A, Ragas P, Lambertsen L, Aaes–Jorgensen A, Molin S, Tolker–Nielsen T (2003). Biofilm formation by Pseudomonas aeruginosa wild type, flagella and type IV pili mutants. Mol. Microbiol. 48 (6): 1511 – 1524.
http://dx.doi.org/10.1046/j.1365-2958.2003.03525.x
PMid:12791135
Korber DR, Lawrence JR, Sutton B, Caldwell DE (1989). Effect of Laminar Flow Velocity on the Kinetics of Surface Recolonization byPseudomonas fluorescens. Microb. Ecol. 18 (1): 1 – 19.
http://dx.doi.org/10.1007/BF02011692
PMid:24196017
Kowalska K, Soscia C, Combe H, Vasseur P, Voulhoux R, Filloux A (2010). The C–terminal amphipathic alpha–helix of Pseudomonas aeruginosa PelC outer membrane protein is required for its function. Biochimie. 92 (1): 33 – 40.
http://dx.doi.org/10.1016/j.biochi.2009.10.004
PMid:19853003
Krogh A, Larsson B, von Heijne G, Sonnhammer EL (2001). Predicting transmembrane protein topology with a hidden Markov model: application to complete genomes. J. Mol. Biol. 305 (3): 567 – 580.
http://dx.doi.org/10.1006/jmbi.2000.4315
PMid:11152613
Lasa I, Penades JR (2006). Bap: a family of surface proteins involved in biofilm formation. Res. Microbiol. 157 (2): 99 – 107.
http://dx.doi.org/10.1016/j.resmic.2005.11.003
PMid:16427771
Lee VT, Matewish JM, Kessler JL, Hyodo M, Hayakawa Y, Lory S (2007). A cyclic–di–GMP receptor required for bacterial exopolysaccharide production. Mol. Microbiol. 65 (6): 1474 – 1484.
http://dx.doi.org/10.1111/j.1365-2958.2007.05879.x
PMid:17824927 PMCid:PMC2170427
Lembre P, Lorentz C, Martino PD (2012). Exopolysaccharides of the Biofilm Matrix: A Complex Biophysical World. In: D. N. Karunaratne (eds), The Complex World of Polysaccharides. InTech Publishers (online open acess). 371 – 392.
http://dx.doi.org/10.5772/51213
Lewandowski Z (2000). Structure and function of biofilms. Biofilms: recent advances in their study and control. L. Evans. Amsterdam, Harwood Academic Publishers. 1 – 17.
López D, Vlamakis H, Kolter R (2010). Biofilms. Cold Spring Harbor Perspectives in Biology 2 (7).
http://dx.doi.org/10.1101/cshperspect.a000398
PMid:20519345 PMCid:PMC2890205
Ma L, Conover M, Lu H, Parsek MR, Bayles K, Wozniak DJ (2009). Assembly and development of the Pseudomonas aeruginosa biofilm matrix. PLoS Pathol. 5 (3): e1000354.
http://dx.doi.org/10.1371/journal.ppat.1000354
PMid:19325879 PMCid:PMC2654510
Ma L, Jackson KD, Landry RM, Parsek MR, Wozniak DJ (2006). Analysis of Pseudomonas aeruginosa conditional psl variants reveals roles for the psl polysaccharide in adhesion and maintaining biofilm structure postattachment. J. Bacteriol. 188 (23): 8213 – 8221.
http://dx.doi.org/10.1128/JB.01202-06
PMid:16980452 PMCid:PMC1698210
Ma L, Lu H, Sprinkle A, Parsek MR, Wozniak DJ (2007). Pseudomonas aeruginosa Psl is a galactose– and mannose–rich exopolysaccharide. J. Bacteriol. 189 (22): 8353 – 8356.
http://dx.doi.org/10.1128/JB.00620-07
PMid:17631634 PMCid:PMC2168683
Ma L, Wang J, Wang S, Anderson EM, Lam JS, Parsek MR, Wozniak DJ (2012). Synthesis of multiple Pseudomonas aeruginosa biofilm matrix exopolysaccharides is post–transcriptionally regulated. Environ. Microbiol. 14 (8): 1995 – 2005.
http://dx.doi.org/10.1111/j.1462-2920.2012.02753.x
PMid:22513190
Macfarlane S, Dillon JF (2007). Microbial biofilms in the human gastrointestinal tract. J. Appl. Microbiol. 102 (5): 1187 – 1196.
http://dx.doi.org/10.1111/j.1365-2672.2007.03287.x
PMid:17448154
Mack D, Fischer W, Krokotsch A, Leopold K, Hartmann R, Egge H, Laufs R (1996). The intercellular adhesin involved in biofilm accumulation of Staphylococcus epidermidis is a linear β–1,6–linked glucosaminoglycan: Purification and structural analysis. J. Bacteriol. 178 (1): 175 – 183.
PMid:8550413 PMCid:PMC177636
Mack D, Nedelmann M, Krokotsch A, Schwarzkopf A, Heesemann J, Laufs R (1994). Characterization of transposon mutants of biofilm–producing Staphylococcus epidermidis impaired in the accumulative phase of biofilm production: Genetic identification of a hexosamine–containing polysaccharide intercellular adhesin. Infect. Immun. 62 (8): 3244 – 3253.
PMid:8039894 PMCid:PMC302952
Maira–Litrán T, Kropec A, Abeygunawardana C, Joyce J, Mark G, Goldmann DA, Pier GB (2002). Immunochemical Properties of the Staphylococcal Poly–N–Acetylglucosamine Surface Polysaccharide. Infect. Immun. 70 (8): 4433 – 4440.
http://dx.doi.org/10.1128/IAI.70.8.4433-4440.2002
PMid:12117954 PMCid:PMC128161
Martínez–Gil M, Yousef–Coronado F, Espinosa–Urgel M (2010). LapF, the second largest Pseudomonas putida protein, contributes to plant root colonization and determines biofilm architecture. Mol. Microbiol. 77 (3): 549 – 561.
http://dx.doi.org/10.1111/j.1365-2958.2010.07249.x
PMid:20545856
McDougald D, Klebensberger J, Tolker–Nielsen T, Webb JS, Conibear T, Rice SA, Kirov SM, Matz C, Kjelleberg S (2008). Pseudomonas aeruginosa : A Model for Biofilm Formation. In: Bernd H.A. Rehm (eds), Pseudomonas : : Model Organism, Pathogen, Cell Factory , Wiley–VCH Verlag GmbH & Co, Weinheim. 215 – 253.
http://dx.doi.org/10.1002/9783527622009.ch9
McDougald D, Rice SA, Barraud N, Steinberg PD, Kjelleberg S (2012). Should we stay or should we go: mechanisms and ecological consequences for biofilm dispersal. Nat. Rev. Microbiol. 10 (1): 39 – 50.
Melchior MB, Vaarkamp H, Fink–Gremmels J (2006). Biofilms: A role in recurrent mastitis infections? Vet. J. 171 (3): 398 – 407.
http://dx.doi.org/10.1016/j.tvjl.2005.01.006
PMid:16624706
Merighi M, Lee VT, Hyodo M, Hayakawa Y, Lory S (2007). The second messenger bis– (3'–5')–cyclic–GMP and its PilZ domain–containing receptor Alg44 are required for alginate biosynthesis in Pseudomonas aeruginosa . Mol. Microbiol. 65 (4): 876 – 895.
http://dx.doi.org/10.1111/j.1365-2958.2007.05817.x
PMid:17645452
Merritt JH, Brothers KM, Kuchma SL, O'Toole GA (2007). SadC reciprocally influences biofilm formation and swarming motility via modulation of exopolysaccharide production and flagellar function. J. Bacteriol. 189 (22): 8154 – 8164.
http://dx.doi.org/10.1128/JB.00585-07
PMid:17586642 PMCid:PMC2168701
Merritt JH, Ha DG, Cowles KN, Lu W, Morales DK, Rabinowitz J, Gitai Z, O'Toole GA (2010). Specific control of Pseudomonas aeruginosa surface–associated behaviors by two c–di–GMP diguanylate cyclases. mBio. 1 (4): 183 – 110.
Mishra M, Byrd MS, Sergeant S, Azad AK, Parsek MR, McPhail L, Schlesinger LS, Wozniak DJ (2012). Pseudomonas aeruginosa Psl polysaccharide reduces neutrophil phagocytosis and the oxidative response by limiting complement–mediated opsonization. Cell. Microbiol. 14 (1): 95 – 106.
http://dx.doi.org/10.1111/j.1462-5822.2011.01704.x
PMid:21951860
Monds RD, Silby MW, Mahanty HK (2001). Expression of the Pho regulon negatively regulates biofilm formation by Pseudomonas aureofaciens PA147–2. Mol. Microbiol. 42 (2): 415 – 426.
http://dx.doi.org/10.1046/j.1365-2958.2001.02641.x
PMid:11703664
Mulcahy H, Charron–Mazenod L, Lewenza S (2008). Extracellular DNA chelates cations and induces antibiotic resistance in Pseudomonas aeruginosa biofilms. PLoS Pathol. 4 (11): e1000213.
http://dx.doi.org/10.1371/journal.ppat.1000213
PMid:19023416 PMCid:PMC2581603
Ochsner UA, Reiser J (1995). Autoinducer–mediated regulation of rhamnolipid biosurfactant synthesis in Pseudomonas aeruginosa . Proc. Natl. Acad. Sci. USA. 92 (14): 6424 – 6428.
http://dx.doi.org/10.1073/pnas.92.14.6424
PMid:7604006 PMCid:PMC41530
Ohman DE, Chakrabarty AM (1982). Utilization of human respiratory secretions by mucoid Pseudomonas aeruginosa of cystic fibrosis origin. Infect. Immun. 37 (2): 662 – 669.
PMid:6811437 PMCid:PMC347583
O'Toole G, Kaplan HB, Kolter R (2000). Biofilm formation as microbial development. Annu. Rev. Microbiol. 54: 49 – 79.
http://dx.doi.org/10.1146/annurev.micro.54.1.49
PMid:11018124
O'Toole GA, Kolter R (1998). Flagellar and twitching motility are necessary for Pseudomonas aeruginosa biofilm development. Mol. Microbiol. 30 (2): 295 – 304.
http://dx.doi.org/10.1046/j.1365-2958.1998.01062.x
PMid:9791175
Overhage J, Schemionek M, Webb JS, Rehm BH (2005). Expression of the psl operon in Pseudomonas aeruginosa PAO1 biofilms: PslA performs an essential function in biofilm formation. Appl. Environ. Microbiol. 71 (8): 4407 – 4413.
http://dx.doi.org/10.1128/AEM.71.8.4407-4413.2005
PMid:16085831 PMCid:PMC1183271
Passador L, Cook JM, Gambello MJ, Rust L, Iglewski BH (1993). Expression of Pseudomonas aeruginosa virulence genes requires cell–to–cell communication. Sci. 260 (5111): 1127 – 1130.
http://dx.doi.org/10.1126/science.8493556
Paul R, Weiser S, Amiot NC, Chan C, Schirmer T, Giese B, Jenal U (2004). Cell cycle–dependent dynamic localization of a bacterial response regulator with a novel di–guanylate cyclase output domain. Genes. Dev. 18 (6): 715 – 727.
http://dx.doi.org/10.1101/gad.289504
PMid:15075296 PMCid:PMC387245
Pearson JP, Gray KM, Passador L, Tucker KD, Eberhard A, Iglewski BH, Greenberg EP (1994). Structure of the autoinducer required for expression of Pseudomonas aeruginosa virulence genes. Proc. Natl. Acad. Sci. USA. 91 (1): 197 – 201.
http://dx.doi.org/10.1073/pnas.91.1.197
PMid:8278364 PMCid:PMC42913
Pedersen SS, Hoiby N, Espersen F, Koch C (1992). Role of alginate in infection with mucoid Pseudomonas aeruginosa in cystic fibrosis. Thorax. 47 (1): 6 – 13.
http://dx.doi.org/10.1136/thx.47.1.6
PMid:1539148 PMCid:PMC463537
Petersen AD, Walker RD, Bowman MM, Schott HC, Rosser EJ (2002). Frequency of Isolation and Antimicrobial Susceptibility Patterns of Staphylococcus intermedius and Pseudomonas aeruginosa Isolates From Canine Skin and Ear Samples Over a 6–Year Period (1992–1997). J. Am. Anim. Hosp. Assoc. 38 (5): 407 – 413.
http://dx.doi.org/10.5326/0380407
PMid:12220023
Petrova OE, Sauer K (2012). Sticky Situations: Key Components That Control Bacterial Surface Attachment. J. Bacteriol. 194 (10): 2413 – 2425.
http://dx.doi.org/10.1128/JB.00003-12
PMid:22389478 PMCid:PMC3347170
Poulsen LV (1999). Microbial Biofilm in Food Processing. LWT – Food Sci. Tech. 32 (6): 321 – 326.
Pratt LA, Kolter R (1999). Genetic analyses of bacterial biofilm formation. Curr. Opin. Microbiol. 2 (6): 598 – 603.
http://dx.doi.org/10.1016/S1369-5274(99)00028-4
Pringle JH, Fletcher M (1983). Influence of substratum wettability on attachment of freshwater bacteria to solid surfaces. Appl. Environ. Microbiol. 45 (3): 811 – 817.
PMid:16346243 PMCid:PMC242376
Prüß B, Verma K, Samanta P, Sule P, Kumar S, Wu J, Christianson D, Horne S, Stafslien S, Wolfe A, Denton A (2010). Environmental and genetic factors that contribute to Escherichia coli K–12 biofilm formation. Arch. Microbiol. 192 (9): 715 – 728.
http://dx.doi.org/10.1007/s00203-010-0599-z
PMid:20559621 PMCid:PMC2923660
Purevdorj–Gage B, Costerton WJ, Stoodley P (2005). Phenotypic differentiation and seeding dispersal in non–mucoid and mucoid Pseudomonas aeruginosa biofilms. Microbiology 151: 1569 – 1576.
http://dx.doi.org/10.1099/mic.0.27536-0
PMid:15870466
Qin Z, Ou Y, Yang L, Zhu Y, Tolker–Nielsen T, Molin S, Qu D (2007). Role of autolysin–mediated DNA release in biofilm formation of Staphylococcus epidermidis. Microbiol. 153: 2083 – 2092.
http://dx.doi.org/10.1099/mic.0.2007/006031-0
PMid:17600053
Rani SA, Pitts B, Beyenal H, Veluchamy RA, Lewandowski Z, Davison WM, Buckingham–Meyer K, Stewart PS (2007). Spatial patterns of DNA replication, protein synthesis, and oxygen concentration within bacterial biofilms reveal diverse physiological states. J. Bacteriol. 189 (11): 4223 – 4233.
http://dx.doi.org/10.1128/JB.00107-07
PMid:17337582 PMCid:PMC1913414
Rocchetta HL, Burrows LL, Lam JS (1999). Genetics of O–antigen biosynthesis in Pseudomonas aeruginosa . Microbiol. Mol. Biol. Rev. 63 (3): 523 – 533.
PMid:10477307 PMCid:PMC103745
Rosenberg M, Bayer EA, Delarea J, Rosenberg E (1982). Role of Thin Fimbriae in Adherence and Growth of Acinetobacter calcoaceticus RAG–1 on Hexadecane. Appl. Environ. Microbiol. 44 (4): 929 – 937.
PMid:16346118 PMCid:PMC242119
Ross P, Mayer R, Benziman M (1991). Cellulose biosynthesis and function in bacteria. Microbiol. Rev. 55 (1): 35 – 58.
http://dx.doi.org/10.1002/cbf.290090304
PMid:1661207
Ross P, Weinhouse H, Aloni Y, Michaeli D, Weinberger–Ohana P, Mayer R, Braun S, de Vroom E, van der Marel GA, van Boom JH, Benziman M (1987). Regulation of cellulose synthesis in Acetobacter xylinum by cyclic diguanylic acid. Nature. 325 (6101): 279 – 281.
http://dx.doi.org/10.1038/325279a0
PMid:18990795
Ruer S, Stender S, Filloux A, de Bentzmann S (2007). Assembly of fimbrial structures in Pseudomonas aeruginosa : functionality and specificity of chaperone–usher machineries. J. Bacteriol. 189 (9): 3547 – 3555.
http://dx.doi.org/10.1128/JB.00093-07
PMid:17293418 PMCid:PMC1855894
Rutherford ST, Bassler BL (2012). Bacterial quorum sensing: its role in virulence and possibilities for its control. Cold Spring Harb Perspect Med. 2 (11).
Ryan RP, Dow JM (2010). Intermolecular interactions between HD–GYP and GGDEF domain proteins mediate virulence–related signal transduction in Xanthomonas campestris. Virulence. 1 (5): 404 – 408.
http://dx.doi.org/10.4161/viru.1.5.12704
PMid:21178479
Ryan RP, Fouhy Y, Lucey JF, Crossman LC, Spiro S, He YW, Zhang LH, Heeb S, Camara M, Williams P, Dow JM (2006). Cell–cell signaling in Xanthomonas campestris involves an HD–GYP domain protein that functions in cyclic di–GMP turnover. Proc. Natl. Acad. Sci. USA. 103 (17): 6712 – 6717.
http://dx.doi.org/10.1073/pnas.0600345103
PMid:16611728 PMCid:PMC1458946
Sa–Correia I, Darzins A, Wang SK, Berry A, Chakrabarty AM (1987). Alginate biosynthetic enzymes in mucoid and nonmucoid Pseudomonas aeruginosa : overproduction of phosphomannose isomerase, phosphomannomutase, and GDP–mannose pyrophosphorylase by overexpression of the phosphomannose isomerase (pmi) gene. J. Bacteriol. 169 (7): 3224 – 3231.
PMid:3036776 PMCid:PMC212373
Sadovskaya I, Vinogradov E, Li J, Jabbouri S (2004). Structural elucidation of the extracellular and cell–wall teichoic acids of Staphylococcus epidermidis RP62A, a reference biofilm–positive strain. Carbohydr. Res. 339 (8): 1467 – 1473.
http://dx.doi.org/10.1016/j.carres.2004.03.017
PMid:15178389
Sakuragi Y, Kolter R (2007). Quorum–sensing regulation of the biofilm matrix genes (pel) of Pseudomonas aeruginosa . J. Bacteriol. 189 (14): 5383 – 5386.
http://dx.doi.org/10.1128/JB.00137-07
PMid:17496081 PMCid:PMC1951888
Samuel G, Reeves P (2003). Biosynthesis of O–antigens: genes and pathways involved in nucleotide sugar precursor synthesis and O–antigen assembly. Carbohydr. Res. 338 (23): 2503 – 2519.
http://dx.doi.org/10.1016/j.carres.2003.07.009
PMid:14670712
Schooling SR, Beveridge TJ (2006). Membrane vesicles: an overlooked component of the matrices of biofilms. J. Bacteriol. 188 (16): 5945 – 5957.
http://dx.doi.org/10.1128/JB.00257-06
PMid:16885463 PMCid:PMC1540058
Sibley CD, Duan K, Fischer C, Parkins MD, Storey DG, Rabin HR, Surette MG (2008). Discerning the complexity of community interactions using a Drosophila model of polymicrobial infections. PLoS Pathol. 4 (10): e1000184.
http://dx.doi.org/10.1371/journal.ppat.1000184
PMid:18949036 PMCid:PMC2566602
Simpson JA, Smith SE, Dean RT (1988). Alginate inhibition of the uptake of Pseudomonas aeruginosa by macrophages. J. Gen. Microbiol. 134 (1): 29 – 36.
PMid:3141564
Simpson JA, Smith SE, Dean RT (1989). Scavenging by alginate of free radicals released by macrophages. Free Radic. Biol. Med. 6 (4): 347 – 353.
http://dx.doi.org/10.1016/0891-5849(89)90078-6
Simpson JA, Smith SE, Dean RT (1993). Alginate may accumulate in cystic fibrosis lung because the enzymatic and free radical capacities of phagocytic cells are inadequate for its degradation. Biochem. Mol. Biol. Int. 30 (6): 1021 – 1034.
PMid:8220249
Smith CS, Hinz A, Bodenmiller D, Larson DE, Brun YV (2003). Identification of Genes Required for Synthesis of the Adhesive Holdfast in Caulobacter crescentus. J. Bacteriol. 185 (4): 1432 – 1442.
http://dx.doi.org/10.1128/JB.185.4.1432-1442.2003
PMid:12562815 PMCid:PMC142846
Stanley NR, Lazazzera BA (2005). Defining the genetic differences between wild and domestic strains of Bacillus subtilis that affect poly–gamma–dl–glutamic acid production and biofilm formation. Mol. Microbiol. 57 (4): 1143 – 1158.
http://dx.doi.org/10.1111/j.1365-2958.2005.04746.x
PMid:16091050
Starkey M, Hickman JH, Ma L, Zhang N, De Long S, Hinz A, Palacios S, Manoil C, Kirisits MJ, Starner TD, Wozniak DJ, Harwood CS, Parsek MR (2009). Pseudomonas aeruginosa rugose small–colony variants have adaptations that likely promote persistence in the cystic fibrosis lung. J. Bacteriol. 191 (11): 3492 – 3503.
http://dx.doi.org/10.1128/JB.00119-09
PMid:19329647 PMCid:PMC2681918
Stewart PS, Franklin MJ (2008). Physiological heterogeneity in biofilms. Nat. Rev. Microbiol. 6 (3): 199 – 210.
http://dx.doi.org/10.1038/nrmicro1838
PMid:18264116
Stoodley P, Dirckx P (2003). Biofilm formation in 3 steps. http://www.biofilm.montana.edu/resources/images/multicellularextracellular/biofilm-formation-3-steps.html. Montana State University, Bozeman, Center for Biofilm Engineering.
Stover CK, Pham XQ, Erwin AL, Mizoguchi SD, Warrener P, Hickey MJ, Brinkman FS, Hufnagle WO, Kowalik DJ, Lagrou M, Garber RL, Goltry L, Tolentino E, Westbrock–Wadman S, Yuan Y, Brody LL, Coulter SN, Folger KR, Kas A, Larbig K, Lim R, Smith K, Spencer D, Wong GK, Wu Z, Paulsen IT, Reizer J, Saier MH, Hancock RE, Lory S, Olson MV (2000). Complete genome sequence of Pseudomonas aeruginosa PAO1, an opportunistic pathogen. Nat. 406 (6799): 959 – 964.
http://dx.doi.org/10.1038/35023079
PMid:10984043
Sutherland IW (2001). Biofilm exopolysaccharides: a strong and sticky framework. Microbiol. 147 (1): 3 – 9.
Sutherland IW (2001). The biofilm matrix–an immobilized but dynamic microbial environment. Trends Microbiol. 9 (5): 222 – 227.
http://dx.doi.org/10.1016/S0966-842X(01)02012-1
Tan MW, Rahme LG, Sternberg JA, Tompkins RG, Ausubel FM (1999). Pseudomonas aeruginosa killing of Caenorhabditis elegans used to identify P. aeruginosa virulence factors. Proc. Natl. Acad. Sci. USA. 96 (5): 2408 – 2413.
http://dx.doi.org/10.1073/pnas.96.5.2408
PMid:10051655 PMCid:PMC26797
Tielker D, Hacker S, Loris R, Strathmann M, Wingender J, Wilhelm S, Rosenau F, Jaeger KE (2005). Pseudomonas aeruginosa lectin LecB is located in the outer membrane and is involved in biofilm formation. Microbiol. 151: 1313 – 1323.
http://dx.doi.org/10.1099/mic.0.27701-0
PMid:15870442
Tolker–Nielsen T, Brinch UC, Ragas PC, Andersen JB, Jacobsen CS, Molin S (2000). Development and dynamics of Pseudomonas sp. biofilms. J. Bacteriol. 182 (22): 6482 – 6489.
http://dx.doi.org/10.1128/JB.182.22.6482-6489.2000
PMid:11053394 PMCid:PMC94796
Tsang PH, Li G, Brun YV, Freund LB, Tang JX (2006). Adhesion of single bacterial cells in the micronewton range. Proc. Natl. Acad. Sci. USA. 103 (15): 5764 – 5768.
http://dx.doi.org/10.1073/pnas.0601705103
PMid:16585522 PMCid:PMC1458647
Ueda A, Wood TK (2009). Connecting quorum sensing, c–di–GMP, pel polysaccharide, and biofilm formation in Pseudomonas aeruginosa through tyrosine phosphatase TpbA (PA3885). PLoS Pathol. 5 (6): e1000483.
http://dx.doi.org/10.1371/journal.ppat.1000483
PMid:19543378 PMCid:PMC2691606
Urch JE, Hurtado–Guerrero R, Brosson D, Liu Z, Eijsink VG, Texier C, van Aalten DM (2009). Structural and functional characterization of a putative polysaccharide deacetylase of the human parasite Encephalitozoon cuniculi. Protein Sci. 18 (6): 1197 – 1209.
http://dx.doi.org/10.1002/pro.128
PMid:19472335 PMCid:PMC2774430
Vallet I, Olson JW, Lory S, Lazdunski A, Filloux A (2001). The chaperone/usher pathways of Pseudomonas aeruginosa : identification of fimbrial gene clusters (cup) and their involvement in biofilm formation. Proc. Natl. Acad. Sci. USA. 98 (12): 6911 – 6916.
http://dx.doi.org/10.1073/pnas.111551898
PMid:11381121 PMCid:PMC34452
Vasseur P, Soscia C, Voulhoux R, Filloux A (2007). PelC is a Pseudomonas aeruginosa outer membrane lipoprotein of the OMA family of proteins involved in exopolysaccharide transport. Biochimie. 89 (8): 903 – 915.
http://dx.doi.org/10.1016/j.biochi.2007.04.002
PMid:17524545
Vasseur P, Vallet–Gely I, Soscia C, Genin S, Filloux A (2005). The pel genes of the Pseudomonas aeruginosa PAK strain are involved at early and late stages of biofilm formation. Microbiol. 151 (Pt 3): 985 – 997.
http://dx.doi.org/10.1099/mic.0.27410-0
PMid:15758243
Venturi V (2006). Regulation of quorum sensing in Pseudomonas . FEMS Microbiol. Rev. 30 (2): 274 – 291.
http://dx.doi.org/10.1111/j.1574-6976.2005.00012.x
PMid:16472307
Wang X, Preston JF, Romeo T (2004). The pgaABCD Locus of Escherichia coli Promotes the Synthesis of a Polysaccharide Adhesin Required for Biofilm Formation. J. Bacteriol. 186 (9): 2724 – 2734.
http://dx.doi.org/10.1128/JB.186.9.2724-2734.2004
PMid:15090514 PMCid:PMC387819
Watanabe M, Sasaki K, Nakashimada Y, Kakizono T, Noparatnaraporn N, Nishio N (1998). Growth and flocculation of a marine photosynthetic bacterium Rhodovulum sp. Appl. Microbiol. Biotechnol. 50 (6): 682 – 691.
http://dx.doi.org/10.1007/s002530051351
Watnick PI, Kolter R (1999). Steps in the development of a Vibrio cholerae El Tor biofilm. Mol. Microbiol. 34 (3): 586 – 595.
http://dx.doi.org/10.1046/j.1365-2958.1999.01624.x
PMid:10564499 PMCid:PMC2860543
Webb JS, Thompson LS, James S, Charlton T, Tolker–Nielsen T, Koch B, Givskov M, Kjelleberg S (2003). Cell death in Pseudomonas aeruginosa biofilm development. J. Bacteriol. 185 (15): 4585 – 4592.
http://dx.doi.org/10.1128/JB.185.15.4585-4592.2003
PMid:12867469 PMCid:PMC165772
West SE, Sample AK, Runyen–Janecky LJ (1994). The vfr gene product, required for Pseudomonas aeruginosa exotoxin A and protease production, belongs to the cyclic AMP receptor protein family. J. Bacteriol. 176 (24): 7532 – 7542.
PMid:8002577 PMCid:PMC197210
Whitchurch CB, Tolker–Nielsen T, Ragas PC, Mattick JS (2002). Extracellular DNA required for bacterial biofilm formation. Science. 295 (5559): 1487.
http://dx.doi.org/10.1126/science.295.5559.1487
PMid:11859186
Whitfield C, Roberts IS (1999). Structure, assembly and regulation of expression of capsules in Escherichia coli. Mol. Microbiol. 31 (5): 1307 – 1319.
http://dx.doi.org/10.1046/j.1365-2958.1999.01276.x
PMid:10200953
Wolfgang MC, Lee VT, Gilmore ME, Lory S (2003). Coordinate regulation of bacterial virulence genes by a novel adenylate cyclase–dependent signaling pathway. Dev. Cell. 4 (2): 253 – 263.
http://dx.doi.org/10.1016/S1534-5807(03)00019-4
Wozniak DJ, Wyckoff TJ, Starkey M, Keyser R, Azadi P, O'Toole GA, Parsek MR (2003). Alginate is not a significant component of the extracellular polysaccharide matrix of PA14 and PAO1 Pseudomonas aeruginosa biofilms. Proc. Natl. Acad. Sci. USA. 100 (13): 7907 – 7912.
http://dx.doi.org/10.1073/pnas.1231792100
PMid:12810959 PMCid:PMC164686
Xu KD, Stewart PS, Xia F, Huang CT, McFeters GA (1998). Spatial physiological heterogeneity in Pseudomonas aeruginosa biofilm is determined by oxygen availability. Appl. Environ. Microbiol. 64 (10): 4035 – 4039.
PMid:9758837 PMCid:PMC106596
Yamanaka S, Watanabe K, Kitamura N, Iguchi M, Mitsuhashi S, Nishi Y, Uryu M (1989). The structure and mechanical properties of sheets prepared from bacterial cellulose. J. Mat. Sci. 24 (9): 3141 – 3145.
http://dx.doi.org/10.1007/BF01139032
Yang L, Barken KB, Skindersoe ME, Christensen AB, Givskov M, Tolker–Nielsen T (2007). Effects of iron on DNA release and biofilm development by Pseudomonas aeruginosa . Microbiol. 153 (5): 1318 – 1328.
http://dx.doi.org/10.1099/mic.0.2006/004911-0
PMid:17464046
Yang L, Hu Y, Liu Y, Zhang J, Ulstrup J, Molin S (2011). Distinct roles of extracellular polymeric substances in Pseudomonas aeruginosa biofilm development. Environ.Microbiol. 13 (7): 1705 – 1717.
http://dx.doi.org/10.1111/j.1462-2920.2011.02503.x
PMid:21605307
Zhang Z, Kulkarni K, Hanrahan SJ, Thompson AJ, Barford D (2010). The APC/C subunit Cdc16/Cut9 is a contiguous tetratricopeptide repeat superhelix with a homo–dimer interface similar to Cdc27. EMBO. J. 29 (21): 3733 – 3744.
http://dx.doi.org/10.1038/emboj.2010.247
PMid:20924356 PMCid:PMC2982764
Zobell CE (1943). The Effect of Solid Surfaces upon Bacterial Activity. J. Bacteriol. 46 (1): 39 – 56.
PMid:16560677 PMCid:PMC373789
Zogaj X, Nimtz M, Rohde M, Bokranz W, Römling U (2001). The multicellular morphotypes of Salmonella typhimurium and Escherichia coli produce cellulose as the second component of the extracellular matrix. Mol. Microbiol. 39 (6): 1452 – 1463.
http://dx.doi.org/10.1046/j.1365-2958.2001.02337.x
PMid:11260463



