Advances in Pharmaceutical and Ethnomedicines
Review Article
Advances in Pharmaceutical and Ethnomedicines 2 (1): 1 – 6Procaspase–3 Activating Therapeutic Agents in Cancer Therapy
Gulzeb Aziz
-
Department of Pharmaceutical Biosciences, School of Pharmacy, University of Oslo, Norway
*Corresponding author:gulzebaziz@gmail.com
ARTICLE CITATION:
Aziz G (2014). Procaspase–3 activating therapeutic agents in cancer therapy. Adv. Pharm. Ethnomed. 2 (1): 1 – 6.
Received: 2014–01–02, Revised: 2014–02–08, Accepted: 2014–02–13
The electronic version of this article is the complete one and can be found online at
(
http://dx.doi.org/10.14737/journal.ape/2014/2.1.1.6
)
which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited
ABSTRACT
Cancer is one of the most deadly groups of diseases faced by human race. The limited success in the treatment of this disease by available anticancer drugs makes it even more challenging and gives the room for new anticancer treatments. Activating apoptosis pathways is a key in cancer chemotherapy. In most of the cancer cells defects in these pathways will result in the resistance to chemotherapy. There are different potential targets in cell death pathways among these the most interesting is procaspase–3 in the recent years. Procaspase–3 cleavage results in the activation of caspase–3 which is the last and decisive player in the esurience of apoptosis. Procaspase activating compound–1 (PAC–1) and 1541 are two very important, potential therapeutic agents which were discovered few years ago. In this review I will discuss in brief about the procaspase–3 as anticancer target as well, will also discuss in detail about PAC–1 and 1541 as activator of this proenzyme. There are quite a number of controversies in the literature about these two potential anticancer therapeutic agents. I will elaborate first anticancer targets inside the cancer cells and then procaspase–3 as potential target and most interestingly PAC–1 and 1541 as potential remedy of cancer. In this review I dug in deep about the future of these two anticancer drugs and also discussed advantages and disadvantages of procaspase–3 as anticancer target and especially, the selectivity of these drugs to target i.e.procaspae–3 which is most important and controversial feature of anticancer therapeutic agents.
Cancer
Cancer is leading cause of death worldwide after cardiovascular and infectious diseases. Cancer is group of diseases which are characterized by alternation in the expression of multiple genes resulting in deregulation of cell division, proliferation and differentiation. The imbalance between cell death and cell proliferation favors the growth of cell population. Malignant tumors have the ability to invade and destroy adjacent tissues and metastasize to other tissues in the body, which is the most important feature of most of the cancers.
Etiology of cancer is complex and relatively unknown in most of the cases but in population certain groups are at risk including smokers, people suffering from infections including HIV, obesity as well people directly or indirectly exposed to the environmental pollutants. In most of the cases cancers are caused by factors described above, ratio of genetically inherited cancers are very low, which is approximately five to ten percent of all the cancers.
In the resent years cancer treatment become a challenge for people working in this field. The most important modes of cancer therapy include radiation, surgery and chemotherapy.
Anticancer Therapies
One of Potential goals in the cancer therapies are maximum effects with minimum side effects which is even more tuff challenge to achieve. All three types of cancer therapies have quit devastating untoward effects. In most of the cases the selectivity and resistance are two important hall marks of the cancer.
Chemotherapy is often used as part of multimodality therapy along with surgery and radiotherapy. Selection of therapy used in order achieve remission is dependent upon tumor type and characteristics. The focus of chemotherapy is historically been achieving increased tumor cytotoxicity based on the difference in proliferation activity between normal and tumor cells. This reflected the classical mode of action of the classical anticancer drugs. Recently, approaches that specifically target the genes, proteins, receptors and molecular pathways that regulate the growth and survival of tumor have gained attention.
The major disadvantages of conversional therapies are selectivity against the cancer cells versus normal cells which result in the potential harmful untoward effects. This continues to inspire research into new anticancer agents. The aim of most these studies are targeted drugs with fewer side effects and increased efficacy which result in a series of novel anticancer drugs.
Cancer chemotherapy is always been challenging for the patients due broad spectrum of unwanted effects and narrow selectivity window against the cancer cells verses normal cells. These issues make even more complications for the researchers in the field of cancer cell biology and drug discovery. Cancer cells are very complex to understand especially mutations and process of resistance towards new chemotherapeutic agents and different cancer types behave differently for the same therapeutic agent and this fact even pose more challenges in this ever changing field.
Potential Anticancer Targets
It is quite interesting to know certain potential targets in cancer cells which are used as war heads to target cancer. In the resent few years many drugs have made and tested against these targets.
Cancer is defined as unchecked growth of the cell and growth cycle is very important steering wheel in the cells and different check points in this cycle are important targets of anticancer agents. The cell cycle consist of distinct phases including G1,S,G2 and M phase as shown in figure 1. In these four phases different check points can be targeted to kill a cancer cell.
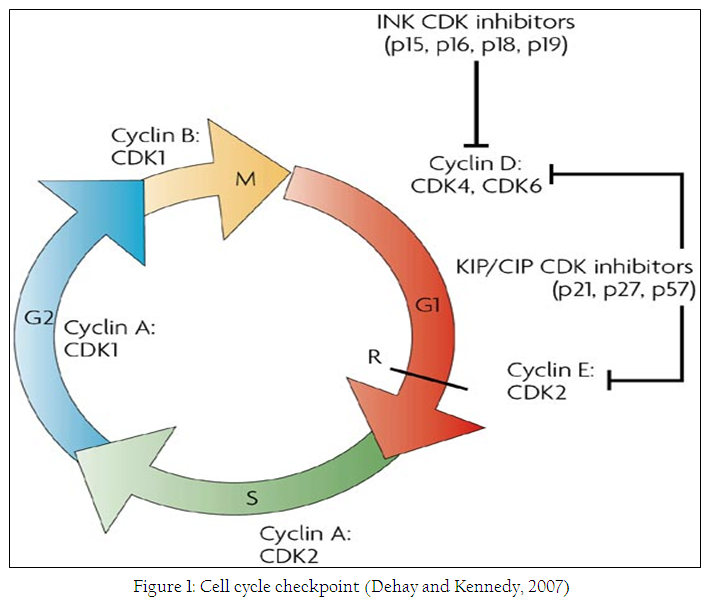
Cell death and apoptosis is another important target in the cancer cells, apoptosis is studied extensively to understand the importance and initiation of this type of cell death in the cancer cells. It is well known fact that most of the cancer cells are resistant to the apoptosis (Peterson et al., 2009a). Apoptosis is also known as programed type of cell death. There are two main pathways that lead to apoptosis in normal and healthy cells including intrinsic and extrinsic pathways. This type of cell death is currently in focus of different research groups around the word. Small molecules to target different proteins including procaspase–3 and IAP’S are studied extensively now a days and one of these small molecules S–PAC–1 is been studied in phase I clinical trials (Peterson et al., 2010).
Cytoskeleton in the cells including actin and tubulin are highly versatile and dynamic structures that determine cell shape and polarity and promote stable cell–cell and cell–matrix adhesion during the morphogenesis (Hall, 2009). These structures are interesting targets in the field of cancer cell biology.
In this review I will discuss in the detail about the procaspase–3 as anticancer drug target due to the fact that one of the small molecule of this class is under phase–1 evaluation and due to certain other controversies around these molecules and procaspase–3 as target (Wolan et al., 2009; Aziz et al., 2010; Boldingh Debernard et al., 2011)
Apoptosis and Cancer
Apoptosis is a programmed multi–step, multi–pathway cell death that occurs in all multicellular organisms. In this process, different biochemical reactions lead to characteristic cell changes and culminate in cell death (Galluzzi et al., 2012). Failure to regulate apoptosis can give rise to several fatal diseases such as autoimmune disorders, neurodegenerative diseases and cancers(Fleischer et al., 2006) For proper and effective treatment of such disorders, it is crucial to understand the molecular mechanisms involved in the process of apoptosis (Mattson, 2000). It is now well documented that most cytotoxic anticancer agents induce apoptosis, raising the intriguing possibility that defects in apoptotic programs contribute to treatment failure (Lowe and Lin, 2000). Resistance to apoptosis is hallmark of most cancer cells which make it even more interesting to study.
Apoptosis is tightly regulated process in the normal cells and the past few year’s researchers in this field has remarkably studied the gene and proteins that are important anticancer targets (Vecchione and Croce, 2010)
The therapeutic goal in cancer treatment is to trigger tumor–selective cell death. However, resistance to treatment is a concern for many types of cancer. Since many anti–neoplastic agents induce an apoptotic type of death in susceptible cells, it is likely that defects or dysregulation of different steps of the apoptotic pathways might be an important determinant of resistance to anticancer drugs. These defects might appear at the initiation and/or execution stages of apoptosis and result in the insufficient elimination of tumor cells, which might lead either to acquired resistance to treatment, or to uncontrolled migration of cancer cells and metastasis. Hence, identification and targeting of the disabled pathway, which is most efficiently inactivated in a particular type of tumor might be the most successful approach in the future. Here we review current knowledge concerning function of apoptotic machinery in cancer cells, and how this information can be used to increase the efficiency of tumor treatment (Rodriguez–Nieto and Zhivotovsky, 2006)
Apoptotic Pathway as Target for Anticancer Drugs
Apoptosis is comprised of intrinsic and extrinsic pathways shown in figure 2. Both intrinsic and extrinsic pathways converge on the activation of procaspase–3 into caspase–3 which is the last executor in the process of apoptosis. If we look in detail the different steps in both of the pathways we will find certain other potential anticancer targets as well but caspase–3 in the last executor in this process and once this is being activated the apoptosis is ensured so it very obvious to target this protein
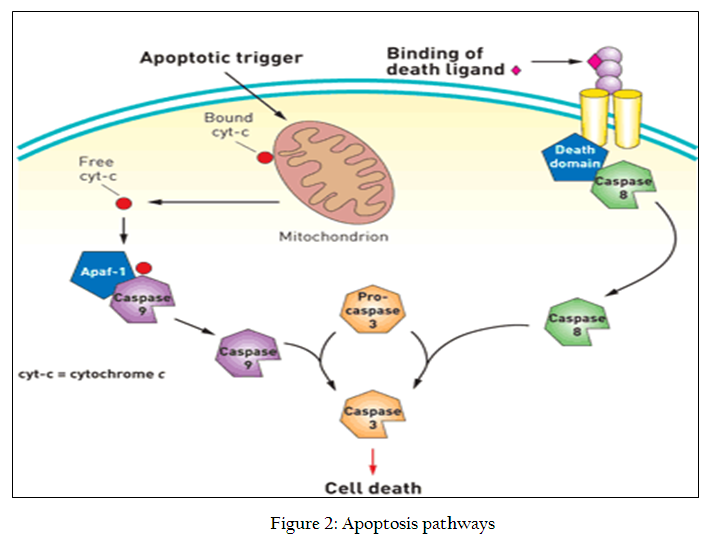
Caspases as Anticancer Target
Caspases are endopeptidases, also called cysteine proteases or thiol proteases are reside inactively nearly all the cells, upon activation they cut hundreds or in some cases thousands of specific proteins. This cleavage leads to the cell death and apoptosis depending upon the function of the caspase.
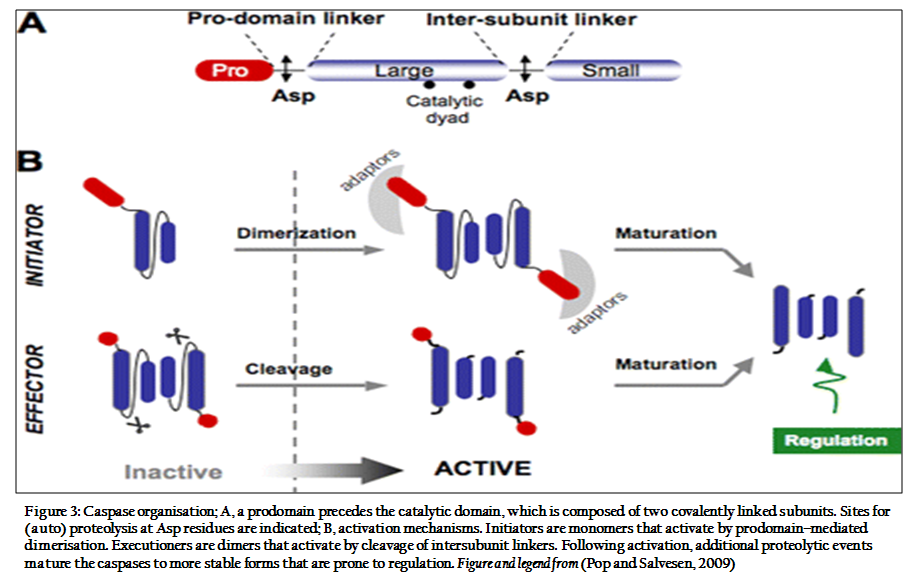
Caspases are classified into two main goups: apoptotic (caspase–2, –3, –6, –7, –8, –9 and –10) and pro–inflamatory (caspase–1, –4, –5, –11, –12 and –13). The apoptotic caspases are further classified into initiators ( caspase–2, –8, –9 and –10) or effectors (caspase–3, –6 and –7)(Pop and Salvesen, 2009)
Caspases are synthesized as inert zymogens that are subsequently activated by internal or external apoptotic stimuli. Once these stimuli occur, cells activate initiator caspases that proteolytically clease and activate effector caspases. Once active this second set of caspases is capable of cleaving a large number of substrates that ultimately lead to cell death.
Procaspases contain an N–terminal prodomain and sequences encoding the large (p20) and small (p10) subunits of procaspas (Kurokawa and Kornbluth, 2009). Caspases specifically recognise a tetrapeptide sequence on their substrate with an absolute requirement of an Asp residue (Vermeulen et al., 2005). The two major apoptotic pathways can be differentiated by the cyt c release from the mitochondria. In the intrinsic pathway, cyt c is released from the mitochondrial intermembrane spaces prior to caspase activation, and in the external pathway, it may be released after the activation of effector caspases by the activation of death receptors (Lossi and Gambino, 2008).
In the both intrinsic and extrinsic apoptotic pathway caspases are placed at key potions, any stimuli either intrinsic or extrinsic may lead to activation of caspase cascade with will ensure the cell death or apoptosis. This unique role makes the caspases as attractive target for anticancer drugs.
PAC–1 as Anticancer Drug
There are many compounds that may initiate apoptosis via the intrinsic or extrinsic pathway in cancer cells and non–cancerous cells. Small molecules are under development to target the apoptotic cascade (Putt et al., 2006). These molecules may be used as anticancer agents or tools to study caspase functions.
Caspase activation by small molecules such as PAC–1 and 1541 has emerged as a powerful anticancer strategy. These molecules are known to transform procaspase–3 (inactive zymogen) to active caspase–3 directly in cancerous cells (Putt et al., 2006), which is a hallmark process of apoptosis.
PAC–1 was the first procaspase–3 activating compound to be discovered by high–throughput screening in 2006 (Putt et al., 2006). It was originally thought that PAC–1 directly transformed procaspase–3 to caspase–3 in vitro and induced apoptosis in cancerous cells isolated from primary colon tumours in a manner directly proportional to the cellular concentration of procaspase–3 (Putt et al., 2006). However, several mechanisms for the activation of caspase–3 by PAC–1 have since been proposed, including the sequestration of inhibitory zinc ions. Zinc is as an endogenous caspase–3 inhibitor and anti–apoptotic agent, which suggests a shared mode of action for certain zinc–chelating anti–cancer compounds (Peterson et al., 2009a),(Peterson et al., 2009b). PAC–1 binds to zinc, enzymatically activating procaspase–3 and allowing the subsequent activation of another molecule of procaspase–3 to form caspase–3 (Peterson et al., 2009a). Zinc chelation is an important mechanism to reduce cell proliferation and induce apoptosis. However, because PAC–1 is not able to fully activate procaspase–3, its potential as an anticancer drug may not be very promising (Wolan et al., 2009).
PAC–1 was found to be neurotoxic in chicken CGN (Aziz et al., 2010) and dogs (Peterson et al., 2010). The neurotoxic properties of PAC–1 raised challenges for its use as an anticancer drug and led to the development of S–PAC–1, which contains a polar sulfonamide on the benzyl ring of PAC–1. This compound is believed to have a similar procaspase–3 activating ability, but because it cannot cross the blood–brain barrier, it is not neurotoxic in mice or dogs (Peterson et al., 2010).
The phenolic hydroxyl group of the PAC–1 molecule is important for caspase activation and zinc chelation. PAC–1 derivatives lacking this group do not activate procaspase–3 in vitro and do not induce death (Peterson et al., 2009b; Aziz et al., 2010). It is also known that ortho–hydroxy N–acyl hydrazones, which are a part of the PAC–1 molecule, bind to metal ions and are essential for zinc binding. Any modification of this structure will result in compounds that are unable to bind zinc or activate caspase–3, which means they would be unable to induce cell death (Peterson et al., 2009b). Conversely, other sites on PAC–1, such as the benzyl ring, could be altered without affecting either activity (Peterson et al., 2009b).
1541 as Anticancer Drug
After the discovery of PAC–1, another small–molecule activator of procaspase–3 was introduced in 2009 and considered as a direct activator. Proenzyme activator 1541 bypasses the upstream pro–apoptotic signalling pathway and induces rapid apoptosis in a variety of cell lines (Wolan et al., 2009). When compared with granzyme B, which is a natural activator of procaspase–3, 1541 induced accelerated activation of caspase–3 to a level that was finally 70% that of granzyme within 2.5 hours. Granzyme B and 1541 both induced full proteolytic cleavage of procaspase–3 into large (17 kD) and small (12 kD) subunits of caspase–3. 1541 promoted the activation and self–processing of procaspase–3 and procaspase–6. 1541 did not activate procaspase–1 and procaspase–7 and was considered to be an excellent activator of procaspase–3 in a variety of cancer cells that are resistant to pro–apoptotic stimuli (Wolan et al., 2009). When compared with PAC–1, 1541 was significantly better at activating procaspase–3 (Wolan et al., 2009).
Which One is Direct Activator: PAC–1 vs 1541
As both the compound PAC–1 and 1541 were claimed as direct activator of procaspase–3 on their introduction in 2006 and 2009 respectively. But these claims are controversial because a study showed that PAC–1 also induces cell death in MCF–7 cells that do not express procaspse–3(Putt et al., 2006; Denault et al., 2007), which indicates additional mechanisms. But EGF and ROS scavenging yielded a small reduction in cell death (Boldingh Debernard et al., 2011). Low concentration of PAC–1 increased the toxicity in PC–12 cells exposed for longer period for the time (Aziz et al., 2010). It is also worth mentioning that PAC–1 and 1541 mediated cell death in PC12 cells when treated with EGF, and these results further augmented by the addition of MEK inhibitor , it showed EGF protects against the cell death through the activation of the MEK/ERK signaling pathway, which regulates transcription or function of apoptotic or antiapototic proteins (Henson and Gibson, 2006). EGF also stimulates the proliferation of PC12 cells through the strong and transient activation of ERK (Sasagawa et al., 2005). These results raise challenges to use of direct activators as anticancer therapeutic agents.
It has been shown that EGF reduces PAC–1 induces caspase–3 activity in PC12 cells. If PAC–1 is a direct activator caspase–3, then caspase–3 depends upon up stream mechanism even when it is activated directly. EGF may up regulate IAP, which directly inhibit caspase–3 and other caspases (Deveraux et al., 1999; Henson and Gibson, 2006). It is also known that EGF can down regulate caspase–9. A third possibility is that PAC–1 does not activate caspase–3 directly, which supported by the cell death in MCF–7 which do not express caspase–3 (Denault et al., 2007). However, 1541 induced cell death was also reduced by EGF, which suggests common mechanism.
It is somewhat difficult to say which one is direct activator of procaspase–3 because both the potential anticancer drugs show similar results.
Conclusion and Future Aspects
In the conclusion, PAC–1 is a zinc chelator in vitro and causes cell death by mechanisms that is partly depends on caspase–3. 1541 seems to have similar effects in cells but has not been proven to be more specific activator of caspase–3 than PAC–1. The crosstalk between the caspase cascade and MEK/ERK–signalling pathway makes it difficult for the compounds to specifically interact with caspase–3. The future of these drugs as potential anticancer therapeutic agents depend upon how these drugs interact with different cancer cell line in vitro and how these drugs behave in vivo when given to cancer patients. I think the most important and concerning challenge is the safety and specifity of these drugs as therapeutic agent which in doubt at the moment.
REFERENCES
Aziz G, Akselsen OW, Hansen T V, Paulsen R E (2010). Procaspase–activating compound 1 induces a caspase–3–dependent cell death in cerebellar granule neurons. Toxicol Appl Pharmacol 247, 238–242.
http://dx.doi.org/10.1016/j.taap.2010.07.002
Boldingh Debernard, K. A., Aziz, G., Gjesvik, A. T., and Paulsen, R. E. (2011). Cell death induced by novel procaspase–3 activators can be reduced by growth factors. Biochem Biophys Res Commun 413, 364–369.
http://dx.doi.org/10.1016/j.bbrc.2011.08.107
Dehay C, and Kennedy H (2007). Cell–cycle control and cortical development. Nat Rev Neurosci 8, 438–450.
http://dx.doi.org/10.1038/nrn2097
Denault, J. B., Drag, M., Salvesen, G. S., Alves, J., Heidt, A. B., Deveraux, Q., and Harris, J. L. (2007). Small molecules not direct activators of caspases. Nat Chem Biol 3, 519; author reply 520.
http://dx.doi.org/10.1038/nchembio0907-519
Deveraux, Q. L., Stennicke, H. R., Salvesen, G. S., and Reed, J. C. (1999). Endogenous inhibitors of caspases. J Clin Immunol 19, 388–398.
http://dx.doi.org/10.1023/A:1020502800208
Fleischer, A., Ghadiri, A., Dessauge, F., Duhamel, M., Rebollo, M. P., Alvarez–Franco, F., and Rebollo, A. (2006). Modulating apoptosis as a target for effective therapy. Mol Immunol 43, 1065–1079.
http://dx.doi.org/10.1016/j.molimm.2005.07.013
Galluzzi, L., Vitale, I., Abrams, J. M., Alnemri, E. S., Baehrecke, E. H., Blagosklonny, M. V., Dawson, T. M., Dawson, V. L., El–Deiry, W. S., Fulda, S., Gottlieb, E., Green, D. R., Hengartner, M. O., Kepp, O., Knight, R. A., Kumar, S., Lipton, S. A., Lu, X., Madeo, F., Malorni, W., Mehlen, P., Nunez, G., Peter, M. E., Piacentini, M., Rubinsztein, D. C., Shi, Y., Simon, H. U., Vandenabeele, P., White, E., Yuan, J., Zhivotovsky, B., Melino, G., and Kroemer, G. (2012). Molecular definitions of cell death subroutines: recommendations of the Nomenclature Committee on Cell Death 2012. Cell Death Differ 19, 107–120.
http://dx.doi.org/10.1038/cdd.2011.96
Hall, A. (2009). The cytoskeleton and cancer. Cancer Metastasis Rev 28, 5–14.
http://dx.doi.org/10.1007/s10555-008-9166-3
Henson, E. S., and Gibson, S. B. (2006). Surviving cell death through epidermal growth factor (EGF) signal transduction pathways: implications for cancer therapy. Cell Signal 18, 2089–2097.
http://dx.doi.org/10.1016/j.cellsig.2006.05.015
Kurokawa, M., and Kornbluth, S. (2009). Caspases and kinases in a death grip. Cell 138, 838–854.
http://dx.doi.org/10.1016/j.cell.2009.08.021
Lossi, L., and Gambino, G. (2008). Apoptosis of the cerebellar neurons. Histol Histopathol 23, 367–380.
Lowe, S. W., and Lin, A. W. (2000). Apoptosis in cancer. Carcinogenesis 21, 485–495.
http://dx.doi.org/10.1093/carcin/21.3.485
Mattson, M. P. (2000). Apoptosis in neurodegenerative disorders. Nat Rev Mol Cell Biol 1, 120–129.
http://dx.doi.org/10.1038/35040009
Peterson, Q. P., Goode, D. R., West, D. C., Ramsey, K. N., Lee, J. J., and Hergenrother, P. J. (2009a). PAC–1 activates procaspase–3 in vitro through relief of zinc–mediated inhibition. J Mol Biol 388, 144–158.
http://dx.doi.org/10.1016/j.jmb.2009.03.003
Peterson, Q. P., Hsu, D. C., Goode, D. R., Novotny, C. J., Totten, R. K., and Hergenrother, P. J. (2009b). Procaspase–3 activation as an anti–cancer strategy: structure–activity relationship of procaspase–activating compound 1 (PAC–1) and its cellular co–localization with caspase–3. Journal of medicinal chemistry 52, 5721–5731.
http://dx.doi.org/10.1021/jm900722z
Peterson, Q. P., Hsu, D. C., Novotny, C. J., West, D. C., Kim, D., Schmit, J. M., Dirikolu, L., Hergenrother, P. J., and Fan, T. M. (2010). Discovery and canine preclinical assessment of a nontoxic procaspase–3–activating compound. Cancer Res 70, 7232–7241.
http://dx.doi.org/10.1158/0008-5472.CAN-10-0766
Pop, C., and Salvesen, G. S. (2009). Human caspases: activation, specificity, and regulation. J Biol Chem 284, 21777–21781.
http://dx.doi.org/10.1074/jbc.R800084200
Putt, K. S., Chen, G. W., Pearson, J. M., Sandhorst, J. S., Hoagland, M. S., Kwon, J. T., Hwang, S. K., Jin, H., Churchwell, M. I., Cho, M. H., Doerge, D. R., Helferich, W. G., and Hergenrother, P. J. (2006). Small–molecule activation of procaspase–3 to caspase–3 as a personalized anticancer strategy. Nat Chem Biol 2, 543–550.
http://dx.doi.org/10.1038/nchembio814
Rodriguez–Nieto, S., and Zhivotovsky, B. (2006). Role of alterations in the apoptotic machinery in sensitivity of cancer cells to treatment. Curr Pharm Des 12, 4411–4425.
http://dx.doi.org/10.2174/138161206779010495
Sasagawa, S., Ozaki, Y., Fujita, K., and Kuroda, S. (2005). Prediction and validation of the distinct dynamics of transient and sustained ERK activation. Nat Cell Biol 7, 365–373.
http://dx.doi.org/10.1038/ncb1233
Vecchione, A., and Croce, C. M. (2010). Apoptomirs: small molecules have gained the license to kill. Endocr Relat Cancer 17, F37–50.
http://dx.doi.org/10.1677/ERC-09-0163
Vermeulen, K., Van Bockstaele, D. R., and Berneman, Z. N. (2005). Apoptosis: mechanisms and relevance in cancer. Ann Hematol 84, 627–639.
http://dx.doi.org/10.1007/s00277-005-1065-x
Wolan, D. W., Zorn, J. A., Gray, D. C., and Wells, J. A. (2009). Small–molecule activators of a proenzyme. Science 326, 853–858.
http://dx.doi.org/10.1126/science.1177585



